Грибы и вызываемые ими болезни
Рассмотрение возбудителей инфекционных болезней начнем с грибов, так как грибы, во-первых, опережают всех остальных паразитов растений по числу патогенных видов, и, во-вторых, вызывают самые опасные и вредоносные болезни.
Американский ботаник Р. Уиттейкер разделил многоклеточные ядерные организмы (эвкариоты) по способу питания на три группы, показанные на схеме:
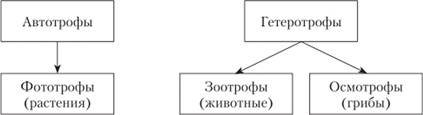
Автотрофами являются организмы, способные создавать сложные органические соединения из неорганических. Растения делают это с помощью солнечной энергии, запасенной в зеленых молекулах хлорофилла, поэтому их называют фототрофами.
Организмы, не способные к синтезу органических веществ из неорганических и нуждающиеся в получении готовых органических соединений извне, называют гетеротрофами. Их Уиттейкер разделил на две группы:
― организмы, поглощающие питательные вещества в виде более мелких организмов или их частей. Внутри их тела поглощенные субстраты с помощью ферментов разлагаются на более простые соединения (перевариваются), которые используются в обмене веществ (метаболизме). Такое питание называют зоотрофным, а обладающих им организмов — животными;
― организмы, поглощающие питательные вещества в виде отдельных молекул из окружающей среды. Поскольку питательные вещества в разных субстратах находятся в виде крупных молекул — полимеров (белков, нуклеиновых кислот, полисахаридов и т.п.), неспособных пройти через клеточные покровы, то эти организмы выделяют ферменты не внутрь, а в окружающее пространство, и разрушают высокомолекулярные полимеры до отдельных "кирпичей", из которых они построены. Эти "кирпичи" всасывают внутрь для использования в метаболизме (внекишечное пищеварение). Такое питание названо осмотрофным, а организмы, обладающие им, — грибами.
Грибы - гетеротрофные эвкариоты, питающиеся осмотрофно.
Способ питания грибов наложил особенности на их строение, физиологию и образ жизни.
Общие свойства грибов
1. Вегетативное тело большинства грибов (мицелий) представляет собой систему разветвленных нитей — гиф, погруженных в субстрат (рис. 1.1). Такое строение при отсутствии специального органа поглощения пищи и всасывания ее всеми клетками тела позволяет максимально оккупировать субстрат.

Рис. 1.1. Грибной мицелий:
а — спора, из которой начался рост мицелия: между гифами наблюдаются многочисленные анастомозы (слияния), армирующие колонию [1]
Но помимо питания грибам необходима адаптация к изменяющимся условиям жизни, вследствие чего из мицелия могут формироваться различные морфологические структуры. Например, для перенесения неблагоприятных погодных условий (летней жары или зимних морозов) грибы образуют покоящиеся структуры. Оболочки отдельных клеток мицелия, а у некоторых грибов — и всех клеток, утолщаются и темнеют, так как в них откладывается темный пигмент — меланин. После этого весь мицелий или участки с такими клетками разваливаются, и образуются покрытые прочными оболочками споры, способные длительный период сохраняться в почве или в растительных остатках. Такие споры называют хламидоспорами. После наступления благоприятных для жизни условий хламидоспоры прорастают ростковой трубкой и формируют новый мицелий. У других видов гифы в отдельных участках мицелия плотно сплетаются, клетки укорачиваются, и образуются клубочки, называемые склероциями. У многих грибов склероции достигают очень больших (конечно, в грибных масштабах) размеров, образуя черные шарики размером с горошину или фиолетовые рожки длиной несколько сантиметров (рис. 1.2). 1 2

Рис. 1.2. Склероции гриба Claviceps purjmrea в колосе ржи [2]
У некоторых грибов черные меланизированные гифы оплетают субстрат, в котором они находятся, превращая его в один большой склероций. Все садоводы знают гниль плодовую яблок, которую вызывают грибы из рода Monilia. Яблоко сначала буреет, его содержимое размягчается, и оно падает с дерева. Еще находясь на ветке или уже упав на землю, оно чернеет, становится жестким и целиком превращается в склероций. В таком виде гриб переносит зимние морозы.
2. Поскольку органические вещества в субстрате находятся в форме полимеров, грибы выделяют внеклеточные ферменты-деполимеразы, расщепляющие полимеры до олиго- и мономеров. Грибы — источники активных деполимераз. Синтез деполимераз, их транспорт через мембраны — это энергоемкие процессы, поэтому в клетках грибов (и бактерий) выработались механизмы, с помощью которых эти ферменты образуются только в нужное время и в нужном месте.
Рассмотрим эти механизмы на примере ферментов, разрушающих пектин (метил-арабиногалактуронан). Этот сложный полимер галактуроновой кислоты входит в состав клеточных стенок растений, а также из него образуются срединные пластинки — межклеточный цемент, склеивающий клетки друг с другом. Грибы образуют набор "отмычек" в виде нескольких ферментов, по-разному разрушающих молекулы пектина. Однако если какой-либо гриб выращивать на искусственной питательной среде, в которую в качестве источника энергии добавлена глюкоза, ни один из этих ферментов синтезироваться не будет. В самом деле, зачем тратить энергию, чтобы разрушать сложные молекулы, освобождать галактуроновые кислоты и превращать их в глюкозу, если в среде есть готовая глюкоза. В природе на всех уровнях жизни господствует принцип экономии.
В случае если вместо глюкозы в качестве единственного источника органических веществ в питательную среду поместить пектин, то в клетках гриба начнут синтезироваться ферменты, разлагающие его. Это явление называют субстратной индукцией, так как субстрат (в данном примере — пектин) сам индуцирует образование фермента, который его разрушает.
Но что будет, если в питательную среду поместить и пектин, и глюкозу? В этом случае, несмотря на наличие субстрата (пектина), ферменты, разрушающие его, формироваться не будут. Согласно принципу экономии, необходимости в этом нет, ведь есть готовая глюкоза. Этот способ генетической регуляции получил название катаболитной репрессии, при которой наличие энергетически более выгодного источника энергии (катаболита) блокирует использование менее выгодного источника.
Денолимеразы грибов широко используются в лесообрабатывающей, легкой и пищевой промышленности, поэтому изучение механизмов генетической регуляции их образования чрезвычайно важно для оптимизации биотехнологических процессов.
3. Грибы накачивают из субстратов в мицелий растворенные моно- и олигомеры благодаря высокому тургорному давлению, развиваемому гифами мицелия. В среднем осмотическое давление, развиваемое грибной гифой, равняется 2,5 атм [3] (равно давлению в хорошо накаченной велосипедной шине). Поэтому клетки всех мицелиальных грибов покрыты прочными оболочками, иначе клетки могли бы лопнуть. Высокое давление необходимо грибам не только для всасывания питательных веществ из окружающей среды, но и для продвижения в плотном субстрате. Ведь грибы развиваются не только в рыхлой почве (хотя глинистые почвы — тоже достаточно плотный субстрат), но и в таких чрезвычайно плотных субстратах, как древесина. Плодовые тела шампиньона, живущего в городских почвах, в процессе развития взламывают асфальт, а кончик гифы многих грибов — паразитов растений разрывает восковой слой на поверхности листьев и наружную стенку эпидермальных клеток.
4. Поскольку вегетативное тело грибов погружено в субстрат, для распространения спор грибы образуют на поверхности субстрата различные органы спороношения (рис. 1.3).
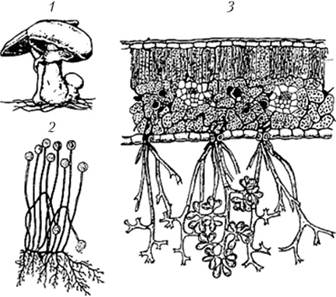
Рис. 1.3. Органы спороношения грибов:
1 — плодовое тело шампиньона, 2 — спорангиеносцы гриба Mucor mucedo с шаровидными спорангиями; 3 — древовидные зооспорангиеносцы возбудителя мильдью винограда оомицета Plasmopara riticola, высовывающиеся из устьиц листа, с лимоновидными зооспорангиями на концах [4]
По своему происхождению спороношения могут быть двух типов половое (телеоморфа) и бесполое (анаморфа). В жизненном цикле большинства грибов они возникают не одновременно, а чередуются друг с другом. У фитонатогенных грибов бесполое спороношение обычно сопровождает паразитическую стадию цикла, а половое — переход в стадию покоя. Так, на листьях и плодах яблони часто встречается заболевание, названное паршой,, при котором возникают темно-оливковые пятна, покрытые бархатистым налетом бесполых спор — конидий. Эти споры, разлетаясь, заражают новые листья и плоды, обеспечивая распространение болезни. А осенью на опавших зараженных листьях происходит половой процесс и формируется зимующая стадия — телеоморфа. Ранней весной половые споры (аскоспоры) осуществляют первичное заражение молодых листьев.
Переход грибов-паразитов из дикорастущих растений на сельскохозяйственные посевы и посадки вызвал у многих видов потерю половой стадии, которая оказалась не только не обязательной, но даже вредной. Например, на родине картофельной фитофторы, в Мексике, паразит зимует в почве и в клубнях диких видов картофеля. Мелкие клубни дикарей в течение теплой мексиканской зимы в массе подвергаются микробному разложению, поэтому более надежный источник сохранения до весны почва, в которой могут сохраняться только половые структуры — ооспоры, покрытые прочными оболочками. Поэтому в дикой природе половой процесс является обязательным. В полях, на которых люди выращивают культурный картофель, бесполые споры паразита, сформированные на листьях, заражают при уборке клубни, и мицелий сохраняется в них до посадки. Поскольку человек создает оптимальные условия, необходимые для сохранности клубней, паразит также получает комфортные условия зимовки и обходится без необходимости тратить энергию на прохождение полового процесса и формирование покоящихся структур (соблюдается принцип максимальной экономии).
У грибов, вызывающих заболевание злаков — ржавчину (название связано с образованием коричневых пятен па листьях и стеблях), летом образуются тонкостенные споры (урединиоспоры), которые разлетаются и заражают новые растения. Осенью, когда листья и стебли начинают высыхать, на соломе формируются толстостенные покоящиеся споры (телиоспоры), которые зимуют. Весной два ядра телиоспоры сливаются, мейотически делятся, и из телиоспоры вырастает промицелий, состоящий из четырех клеток (базидия), на которой образуются четыре гаплоидные споры. Эти споры не способны заразить листья ржи или пшеницы, они должны заразить листья так называемого промежуточного хозяина, разного для разных видов ржавчины, на котором пройдет половой процесс. И только образующиеся в результате полового процесса споры (эциоспоры) заражают злаки.
Подобная стратегия выработалась в дикой природе, где злаковые травы росли в окружении других растений, а фитоценозы (сообщества растений) были очень сложными, многочленными. Но человек начал возделывать пшеницу, ячмень или овес на огромных полях, стал уничтожать сорные и другие сопутствующие растения, так что вероятность попадания гаплоидных базидиоспор на листья промежуточного хозяина стала проблематичной. К тому же крестьяне придумали во многих зонах, где зимы не слишком суровы, высевать семена пшеницы и ржи не весной, а в конце лета, чтобы всходы сохранялись под снегом и на следующий год давали урожай более ранний и более высокий (озимые культуры). Этим прекрасно воспользовались паразиты: их летние споры (урединиоспоры) перелетали со стареющих растений посева прошлого года на новые всходы и зимовали в их листьях под снегом. Образование толстостенных телиоспор, формирование промицелия, обязательное попадание гаплоидных базидиоспор на листья промежуточного хозяина и половой процесс на его листьях
оказались ненужными. Жизненный цикл, опять же вследствие принципа максимальной экономии, чрезвычайно упростился, зато численность урединиоспор значительно выросла вследствие той же экономии энергии.