Транспирация ипередвижение соды в листьях
Передвижение воды у цветковых астений
Вода, находящаяся в растении, непосредственно контактирует сводой почвы и с парами воды в окружающем растение воздухе. Мы уже говорили о том, что вода движется от более высокого водного потенциала к более низкому. Поэтому, как полагают физиологи растений, вода передвигается по растению из области с высоким водным потенциалом (т.е. из почвы) в область с более низким водным потенциалом (т.е. в атмосферу) в соответствии с градиентом водного потенциала. Водный потенциал в умеренно влажном воздухе на несколько десятков тысяч килопаскалей ниже, чем в растении; отсюда и большее стремление воды выходить из растения.
Основная масса воды поступает в растение через корневые волоски. Вода проходит через кору корня в ксилему, поднимается по ксилеме к листьям и испаряется с поверхности клеток мезофилла, а затем диффундирует наружу через устьица. Последний процесс называется транспирацией,а поток воды из корня к транспирирующей поверхности-транспира-ционным током.Установлено, что растение в среднем использует менее 1% поглощаемой им воды. Куда расходуется вода, мы уже рассматривали в разд. 5.1.2.
Транспирация ипередвижение соды в листьях
В обычных условиях вода выходит из растения в виде пара. Для перехода воды из жидкого состояния в парообразное нужна дополнительная энергия, которую называют скрытой теплотой парообразования.Ее дает солнце (солнечная энергия), и согласно наиболее принятой теории транспирации, теории сцепления-натяжения,именно эта энергия обеспечивает общий поток воды через растение. Транспирация - это испарение воды с поверхности растения; ниже перечислены три основные структуры, участвующие в этом испарении (в скобках указана относительная потеря воды через них при открытых устьицах).
1. Устьица- поры, через которые диффундирует вода, испарившаяся с поверхности клеток; такие поры есть в эпидермисе листьев и зеленых стеблей (около 90% воды).
2. Кутикула- восковой слой, покрывающий эпидермис листьев и стеблей; через него проходит вода, испаряющаяся с наружных стенок клеток эпидермиса (около 10%; больше или меньше в зависимости от толщины кутикулы).
3. Чечевички:обычно их роль в испарении воды очень мала, но у листопадных деревьев после сбрасывания листвы через чечевички теряется основная масса воды.
Потери воды в результате транспирации могут быть очень велики. Травянистые растения, например хлопчатник или подсолнечник, ежедневно теряют до 1-2 л воды, а старый дуб-более 600 л.
Почему транспирация идет главным образом через листья?
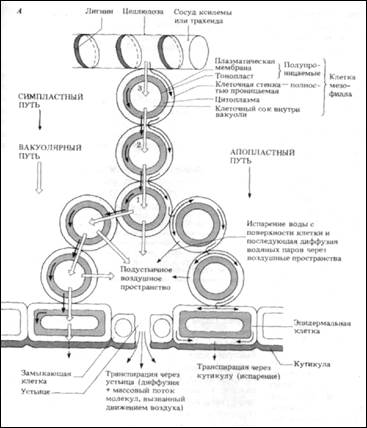
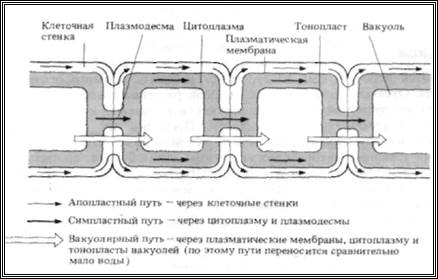
Вода поступает в листья по ксилеме. В листьях есть своя собственная, сильно разветвленная сеть проводящих пучков, охватывающая весь лист. Ее ветви оканчиваются одним или несколькими сосудами или трахеидами, которые почти не лиг-нифицированы. Поэтому вода легко проходит через их целлюлозные стенки и поступает к снабжаемым ими клеткам мезофилла. На рис. 14.7 показаны три основныхпути, по которым вода идет дальше: апопластный (через клеточные стенки), симпласт- I ный (через цитоплазму и плазмодесмы) и вакуо-лярный (от вакуоли к вакуоли).
14.3.1. Путь через апопласт
Апопласт-этосистема, образованная из примыкающих друг к другу клеточных стенок, создающих непрерывное целое (за исключением поясков Каспари в корнях, разд. 14.5.2). «Свободное пространство», которое может быть заполнено водой, занимает до 50% объема клеточных стенок. Испарение воды из апопласта мезофилла в межклеточное воздушное пространство создает натяжение в непрерывном потоке воды через апопласт, и вода массовым потоком проходит через стенки в результате сцепления (когезии) ее молекул (разд. 14.4). В апопласт вода поступает из ксилемы.
14.3.2. Путь через симпласт
Симпласт-это система связанных между собой протопластов растения. Цитоплазма соседних клеток соединена плазмодесмами-цитоплазма-тическими тяжами, проходящими через поры в примыкающих друг к другу клеточных стенках (рис. 14.1, Б). Детали строения плазмодесм до сих пор окончательно не выяснены, поэтому не известно, в какой мере они служат каналами для передвижения веществ. Кажется, однако, вероятным, что если уж вода и растворенные в ней вещества проникнут в цитоплазму одной из клеток, то дальше они могут переходить из клетки в клетку, минуя мембраны. Как и при вакуолярном переносе (разд. 14.3.3), вода идет по градиенту водного потенциала. Возможно, что этому способствуют и токи цитоплазмы. Передвижение воды по симпласту имеет большее значение, чем путь через вакуоли.
14.3.3. Вакуолярный путь
В этом случае вода переходит из вакуоли в вакуоль через другие компоненты смежных клеток;
при этом она пересекает симпласт и апопласт и проходит через плазмалемму и тонопласт путем осмоса (рис. 14.7, Б). Она движется по градиенту водного потенциала, который создается как описано ниже.
Вода испаряется с увлажненной поверхности клеточных стенок мезофилла и попадает в межклетники, в частности в околоустьичиые воздушные пространства. Рассмотрим, например, клетку 1 на рис. 14.7, А. Потеря ею воды приводит к тому, что в ней падает тургорное давление и понижается водный потенциал. В результате этого водный потенциал клетки 2 становится выше, чем у клетки 1 (хотя в состоянии равновесия он был одинаков). Поэтому вода идет из клетки 2 в клетку 1, и водный потенциал клетки 2 понижается по сравнению с потенциалом клетки 3. Так в листе создается градиент водного потенциала на всем пути от сосудов ксилемы с ее более высоким водным потенциалом до клеток мезофилла, потенциал которых гораздо ниже. За счет осмоса вода поступает из ксилемы в клетки мезофилла. Хотя нам удобнее рассматривать передвижение воды шаг за шагом, следует подчеркнуть, что градиент водного потенциала, который создается в листе, на самом деле непрерывен, и вода равномерно течет по градиенту, подобно тому как жидкость движется по фитилю.
Иногда создается неверное представление, что вода передвигается в листе под действием градиента осмотического давления. Однако нет никаких данных, которые бы позволили предположить, что осмотическое давление в соответствующих клетках сильно различается, хотя градиент водного потенциала, несомненно, существует. Разность водного потенциала возникает в основном из-за разности тургорного давления (вспомните, что потеря даже небольшого количества воды в первую очередь влияет на тургорное давление, а не на осмотическое). То же самое можно сказать и о корне (разд. 14.5), в котором есть градиенты тургорного давления и водного потенциала, но не всегда существуют градиенты осмотического давления.
14.3.4. Выход воды через устьица
Все три пути переноса воды завершаются испарением ее в воздушные пространства листа. Отсюда пары воды диффундируют через устьица по пути наименьшего сопротивления, т. е. от более высокого водного потенциала внутри листа к более низкому за его пределами. У двудольных растений устьица обычно располагаются в основном на нижней поверхности листа, на верхней же их меньше или нет вовсе. Регуляцию открывания и закрывания устьиц мы рассмотрим в разд. 14.3.9. Все листья окружены неподвижным слоем воздуха, толщина которого определяется размерами листа, особенностями его строения, например опущенностью, и скоростью ветра. Пары воды сначала диффундируют через этот слой и лишь затем уносятся воздушными потоками. Чем тоньше неподвижный слой, тем быстрее идет транспирация. Существует диффузионный градиент, направленный от неподвижного слоя назад к клеткам мезофилла. Теоретически каждое устьице должно иметь свой собственный диффузионный градиент-свою «диффузионную оболочку» (рис. 14.8). На самом же деле, если воздух неподвижен, диффузионные оболочки соседних устьиц перекрываются и образуют одну общую оболочку.