Природа бактерій, фагів та їх застосування на практиці
Припущення, що бактеріофаги мають корпускулярну природу, було висунуто ще Ф.д'Ерелєм. Однак тільки після винайдення електронного мікроскопа вдалося побачити і вивчити ультраструктуру фагів. Нагадаємо, що довгий час уявлення про морфологію та основні особливості фагів грунтувалися на результатах вивчення фагів Т-групи — ТІ, Т2, ., ТІ, які розмножуються на Е.соli штаму В. Однак з кожним роком з'являлися нові дані щодо морфології і структури різноманітних фагів, що зумовило необхідність їхньої морфологічної класифікації.
Вивчення хімічного складу фагів показало, що він досить простий; по суті фаги є нуклеопротещами, тобто складаються в основному з білка і нуклеїнової кислоти. Фагові частинки мають кілька різних білків, насамперед структурних, які становлять капсид головки і елементи відростка (чохол, стрижень, базальну пластинку і нитки). У головці булавоподібних фагів є також внутрішній білок (3-7 % загального вмісту білка). У фагів виявлено ферменти лізоцим, фосфатазу та деякі інші. Білки виконують різні функції: захищають нуклеїнову кислоту від пошкоджень і дії ферментів нуклеаз, беруть участь у тісному контакті фага з бактеріальною клітиною, забезпечують через ферментативну дію процес зараження тощо.
Другою важливою складовою частиною фагів є нуклеїнові кислоти. У фагів, як і в інших вірусів, є тільки один тип нуклеїнової кислоти — ДНК або РНК. Цією властивістю віруси відрізняються від інших мікроорганізмів, в клітинах яких є обидва типи нуклеїнових кислот. У фагів виявлено дволанцюгову ДНК (найчастіше) і одно-ланцюгові ДНК та РНК. Залежно від типу своєї нуклеїнової кислоти фаги поділяють на ДНК-вмісні і РНК-вмісні. Нуклеїнова кислота щільно упакована у головці фага.
У деяких фагів знайдено невеличку кількість ліпідів (2,5-10,5 %), переважно жирних кислот і фосфоліпідів, а також сліди вуглеводів. Значення цих компонентів поки що недостатньо вивчено. Вважають, що ліпіди та інші компоненти (подібно до інших вірусів) мають клітинне походження і фаговий геном не кодує їхнього синтезу.
Бактеріофаги володіють антигенними властивостями. При багаторазовому парентеральному введенні фагів кролям або іншим тваринам можна одержати сироватки, які містять сферичні антитіла до відповідних фагів. Такі сироватки називають антифаговими. Антитіла таких сироваток здатні давати з відповідними фагами звичайні серологічні реакції — аглютинації, преципітації, зв'язування комплемента, а також спричинюють нейтралізацію літичної активності фагів. Антифагові сироватки строго специфічні. Цю властивість часто використовують при серологічній класифікації фагів.
Уперше різні фаги застосували Ф. д'Ерель та інші дослідники, вивчаючи кишкові інфекції у людини. Практикувалось використання фагів також у хірургічній та акушерсько-гінекологічній практиці при інфекційних процесах, які спричинюються стафілококами, анаеробними клостридіями, а також
в офтальмології і стоматології. Поряд з цим нагромадилось багато даних про відсутність лікувального ефекту при застосуванні фагів. Вважають, що основною причиною цього є невдалий підбір фагів. Широке впровадження в практику сульфаніламідних препаратів і антибіотиків також зменшило інтерес до питань фагопрофілактики і фаготерапії бактеріальних інфекцій.
Фагопрофілактика і фаготерапія застосовуються і при різних захворюваннях тварин. При паратифі телят використовують гертнер-фаг, при захворюванні поросят — суйпестифер-фаг, при колібактеріозі — колі-фаг та інші.
Бактеріофаги черевнотифозної сальмонели, кишкової палички та інших бактерій можуть міститися у воді річок, ставків, водосховищ, колодязів і в забруднених стічних водах. Виявлення фагів у воді або в грунті може бути показником забруднення цих середовищ відповідними бактеріями. Наприклад, виявлення коліфага є показником фекального забруднення води.
Поряд з цим слід зазначити, що фаги нерідко можуть завдавати й великої шкоди, зокрема при виробництві антибіотиків, молочнокислих продуктів, бактеріальних добрив, гальмуючи розвиток корисних мікробів. Зараження фагами бульбочкових бактерій, азотобактера та інших азотфіксаторів призводить до затримки їхнього розвитку, а це погіршує процес фіксації молекулярного азоту, а отже, негативно впливає на врожай.
Плазміди бактерій
Відомо, що ядра в еукаріотних клітинах, а нуклеоїди в прокаріот-них є місцем локалізації генетичного матеріалу. Генетичний апарат у бактерій складається з молекули ДНК, замкненої в кільце. Довжина кільця може сягати 1,0—1,4 мм. Воно міститься в нуклеарній ділянці бактеріальної клітини. Гігантська кільцева молекула ДНК, яка складається із функціонально неоднорідних генетичних детермінант генів, дістала назву бактеріальної хромосоми.
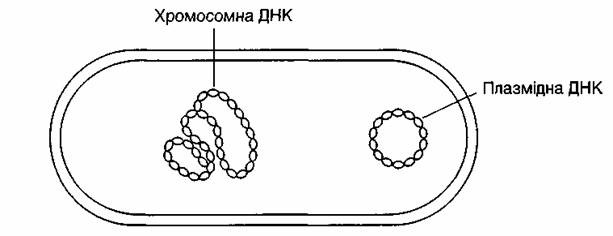
Рис. 1