А передня група ядер (передній гіпоталамус)регулює функції парасимпатичної нервової системи
Ушкодження або руйнування різних відділів гіпоталамуса спричинюють зміни відповідно до їх функціонального призначення: афагії (відмова від їжі), смерті від голоду, гіпоталамічну гіперфагію (жадобу до їжі).
-захисна поведінкова реакція, яка має назву несправжньої люті. У людей такий тип поведінки вважається емоційним виразом страху і люті.
- знаходиться також центр захисної поведінки (неподалік від харчового). Стимуляція цієї зони зумовлює у ссавців чіткі захисні поведінкові реакції, які мають назву несправжньої люті, оскільки вони, як правило, безадресні. Такі реакції є видоспецифічними. Вони супроводжуються слиновиділенням, розширенням зіниць, пілоерекцією, почастішанням дихання. У людей такий тип поведінки вважається емоційним виразом страху і люті.
До моменту народження більша частина ядер згір'я добре розвинена. Після народження відбувається збільшення згір'я. Цей процес триває близько 13-15 років. Ядра закінчують свій розвиток в період статевого дозрівання.

Функції спинного мозку
Спинний мозок виконує дві основні функції: рефлекторну і провідну.
Рефлекторна функція спинного мозку полягає у здійсненні складних рухових реакцій організму. -Спинний мозок іннервує всю скелетну мускулатуру, крім м'язів голови, які іннервуються черепними нервами.
-У спинному мозку містяться рефлекторні центри мускулатури тулуба, кінцівок шиї.
- Тут же розташовано багато центрів автономної нервової системи.
-Рефлекси сечовипускання і дефекації зв'язані з функцією спинного мозку.
Соматичні рефлекси - це рухові рефлекси кінцівок, тулуба чи його частин.
Рефлекси розтягання, або власні міотатичні рефлекси м'язів, - це рефлекси, які ініціюються подразненням пропріорецепторів - рецепторів скелетних м'язів або їхніх сухожилків.
Сухожильні рефлекси належать до числа рефлексів, що мають велике значення для підтримування положення тіла, швидкого встановлення рівноваги. Вони виконуються дуже швидко, їх рефлекторна дуга має мінімальну кількість елементів - це двонейронна дуга. Рецептори сухожильних рефлексів знаходяться у скелетних м'язах (пропріоцептори) і збуджуються при розтягуванні м'язових волокон. Якщо вдарити по сухожиллю, то м'яз розтягується і це приводить до збудження рецепторів і виникнення рефлексу.
Сухожильні рефлекси - колінний, п'ятковий - завжди можна викликати у дитини на першому ж році життя. Формування їхніх рефлекторних дуг відбувається ще на 6-7-му місяці антенатального розвитку.
Колінний рефлекс у немовлят супроводжується скороченням привідних м'язів протилежної ноги, внаслідок чого нога повертається всередину. Цю реакцію називають перехресним рефлексом привідних м'язів. Колінний рефлекс у дітей не спостерігається після 7 місяців, тому що його гальмують вищі центри нервової системи. Знову у дітей з'являється колінний рефлекс після 2 років і надалі зберігається на все життя. П'ятковий рефлекс з'являється у більшості дітей після 7-8 місяців і також існує протягом всього життя.
Зникнення або зміна сили сухожильних рефлексів свідчать про несприятливі зміни в здоров'ї дитини. Визначення сухожильних рефлексів має велике значення для перевірки функціонального стану організму в цілому та нервової системи зокрема.
Крім рухових, спинний мозок здійснює велику кількість вісцеральних рефлексів, за допомогою яких регулює функцію внутрішніх органів і систем. Звичайно, ці рефлекси перебувають під контролем вищих відділів ЦНС, зокрема, кори великого мозку.
Спинний мозок здійснює і провідну функцію. Доцентрові імпульси, які надходять у спинний мозок по задніх корінцях, передаються по провідних шляхах спинного мозку у відділи головного мозку, що лежать вище. У свою чергу, із відділів центральної нервової системи спинний мозок одержує імпульси, які можуть змінювати діяльність скелетної мускулатури і внутрішніх органів. Провідні шляхи діляться на висхідні і низхідні. Висхідні шляхи проводять імпульси із периферії (від рецепторів шкіри, м'язів, суглобів, внутрішніх органів) до головного мозку. Низхідні шляхи проходять по передніх і частково бічних стовпах. По них передаються імпульси від головного мозку до рухових центрів спинного мозку, а від них ці імпульси йдуть до скелетних м'язів.
Розвиток спинного мозку
Коли в ембріона головний мозок знаходиться на стадії мозкових пухирів, спинний мозок досягає значних розмірів. На ранніх стадіях розвитку плоду спинний мозок заповнює всю порожнину хребетного каналу. Потім хребетний стовп випереджає в рості спинний мозок і до моменту народження закінчується на рівні III поперекового хребця. У новонароджених довжина спинного мозку - 14-16 см - до 10 років подвоюється. На поперековому зрізі спинного мозку дітей раннього віку відмічається перевага передніх рогів над задніми. Провідні шляхи досить розвинені вже на момент народження. Мієлінізація їх волокон закінчується до 3-х місяців, тоді як у периферичних нервах тягнеться до 3-х років і більше.
Із збільшенням маси спинного мозку змінюється і його довжина, яка у чоловіків досягає 46 см, а у жінок 41 - 42 см. Під час розвитку спинний мозок не встигає за ростом хребта, і тому у дитини спинний мозок виповнює лише дві третини хребтового каналу.
Функції варолієвого моста
Вароліїв міст,або міст (pons), розташований на нижній поверхні головного мозку у вигляді широкого виступу з поперечною посмугованістю (рис. 43, 48). Він межує спереду з ніжками великого мозку, позаду - з довгастим мозком, а по боках переходить у середні мозочкові ніжки, в товщі яких виступають корінці трійчастого нерва. Між мостом і пірамідою виходить відвідний нерв, а позаду і збоку лицевий і присінково-завитковий.
Основна частина моста складається переважно з поздовжніх і поперекових волокон моста.
Поздовжні волокна утворені кірково-мостовими волокнами, що розташовані зовні і зв'язують міст з корою великого мозку, а також волокнами пірамідного шляху. Поздовжні волокна діляться на кірково-ядерні, які йдуть від кори великого мозку до рухових ядер черепних нервів протилежного боку; кірково-спинно-мозкові, що прямують до рухових клітин передніх рогів сірої речовини спинного мозку протилежного боку, та кірково-сітчасті волокна - від кори головного мозку.
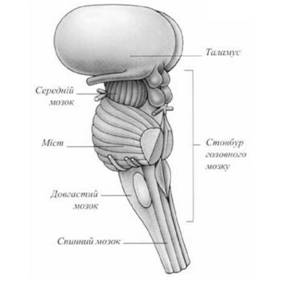
Рис. 48. Таламус і стовбур головного мозку (за Дорлінг Кіндерслі, 2003)
Поперекові волокна моста, які становлять основну масу передньої частини, зв'язують міст із мозочком, йдучи в складі середніх мозочкових ніжок.
У задній частині - покриві моста - закладені утвори:
o сітчастий, що є продовженням сітчастого утвору довгастого мозку;
o ядра п'яти черепних нервів;
o волокна спинно- і середню мозкового шляхів трійчастого нерва з їхнім перехрестям;
o низхідні волокна червоно-ядерно-спинномозкового, сітчасто-спинномозкового шляхів, які закінчуються на рухових клітинах передніх рогів сірої речовини спинного мозку.
Ядра моста беруть участь у регуляції сльозові потовиділення, координації актів жування і ковтання, кліпання очима тощо. Парасимпатичне ядро блукаючого нерва входить до системи рефлекторної регуляції дихання і кровообігу.
Будова і функції мозочка
Мозочок розташований за довгастим мозком і варолієвим мостом (рис. 43), має дві півкулі, з'єднані черв'ячком. Сіра речовина мозочка утворює кору. Поверхня мозочка поділена поперечними борознами. Біла речовина знаходиться під корою, всередині якої міститься чотири ядра сірої речовини.

Рис. 49. Найважливіші міжнейронні зв'язки мозочка (за Чайченко П. М., Цибенко В. О., Сокун В. Д., 2003):
А - молекулярний шар; Б - шар грушоподібних нейронів;
В - зернистий шар; Г - ядра мозочка; 1 - зірчастий нейрон;
2 - кошиковий нейрон; 3 - грушоподібний нейрон (клітина Пуркіньє);
4 - великий зірчастий нейрон (клітина Гольджі);
5 - зерноподібний нейрон; 6 - мохоподібні нервові волокна;
7 - висхідне нервове волокно (ліаноподібне);
8 - паралельні нервові волокна
Сіра речовина - кора мозочка - має три шари (рис. 49): поверхневий молекулярний шар, шар грушоподібних нейронів (клітин Пуркін'є) і нижній шар - зернистий.
У шарах кори мозочка розміщені шість різних типів нейронів. Зерноподібні нейрони, кількість яких у людини становить 1010 - 1011, розміщені у зернистому шарі, а їхні аксони прямують до молекулярного шару, де розгалужуються Т-подібно, посилаючи в обох напрямках уздовж поверхні кори гілочки (паралельні нервові волокна). Ці волокна проходять через ділянки розгалуження дендритів інших п'яти типів нейронів і утворюють на них синапси. У зернистому шарі розміщені великі зірчасті нейрони (клітини Гольджі), дендрити яких поширюються на відносно невелику відстань, а аксони прямують до зерноподібних клітин.
Шар грушоподібних нейронів налічує у людини близько 15 млн. нейронів, дендрити яких розгалужуються у молекулярному шарі. Аксони грушоподібних нейронів спускаються до ядер мозочка, а невелика їх кількість закінчується на пристінкових ядрах. Аксони останніх клітин становлять єдиний еферентний шлях від мозочка.
Зв'язок мозочка з іншими структурами мозку здійснюється за допомогою трьох пар ніжок. Нижні ніжки зв'язують мозочок зі спинним і довгастим мозком, найтовщі середні ніжки охоплюють довгастий мозок і, розширюючись, переходять на міст, що сполучає мозочок з корою великого мозку, а через верхні ніжки проходять еферентні та аферентні шляхи.
Мозочок сполучається з довгастим мозком, варолієвим мостом і середнім мозком за допомогою численних нервових волокон, які утворюють три пари ніжок мозочка.
Останні три шари нейронів кошикові і зірчасті нейрони та клітини Лугаролежать у молекулярному шарі. Аксони кошикових нейронів закінчуються на тілі, а зірчастих - на дендритах грушоподібних нейронів.
До кори мозочка входять два типи волокон. Висхідні нервові волокна (ліаноподібні) проходять крізь зернистий шар і закінчуються у молекулярному на дендритах грушоподібних нейронів, а дуже численні (у людини - до 50 млн.) мохоподібні волокна закінчуються на зірчастих нейронах. До кожного нейрона кори мозочка підходять численні паралельні нервові волокна від зірчастих клітин, і тому через ці нейрони на будь-якій клітині кори мозочка конвертують (сходяться) сотні мохоподібних волокон.
Висхідні (ліаноподібні) волокна утворюють численні збуджувальні синапси на дендритах грушоподібних нейронів, і тому досить одного стимулу, щоб грушоподібний нейрон відповів цілим розрядом імпульсів. Мохоподібні волокна збуджують зірчасті нейрони, які через паралельні нервові волокна виявляють збуджувальний вплив на решту нейронів. Проте всі ці нейрони є гальмівними: великі зірчасті нейрони гальмують зірчасті нейрони, а розряди грушоподібних нейронів гальмують нейрони ядер мозочка. Отже, за винятком зірчастих, усі нейрони кори мозочка виконують гальмівні функції.
Грушоподібним нейронам властива спонтанна імпульсна активність у стані спокою, чим і зумовлюється тонічне гальмування нейронів ядер мозочка. Зі зростанням активності цих гальмівних грушоподібних нейронів завдяки збудженню мохоподібних чи висхідних волокон гальмування нейронів ядер мозочка значно підсилюється. Якщо ж грушоподібні нейрони гальмуються (безпосередньо-зірчастими або кошиковими нейронами, опосередковано за допомогою великих зірчастих нейронів), то це призводить до розгальмування структур мозочка, розміщених нижче.
Аксони грушоподібних нейронів закінчуються переважно на нейронах підкіркових мозочкових ядер (найбільше з них - зубчасте, ядро вершини (фастигіальне), коркоподібне й кулясте (інтерпозитні)). Нейрони від цих ядер прямують до стовбура головного мозку. Коркоподібне й кулясте ядра відповідають за підтримання пози тіла, а зубчасте - за точність, швидкість і плавність рухів. Проте цілісна свідома і несвідома рухова функція здійснюється завдяки постійній взаємодії мозочкових і позамозочкових структур.
До мозочка надходять імпульси від усіх рецепторів, які подразнюються під час рухів тіла. Мозочок бере участь у координації складних рухових актів. Двобічні зв'язки мозочка і кори великого мозку дають йому змогу впливати на довільні рухи.
Мозочок впливає на функцію деяких автономних центрів, проте головна його роль - це забезпечення узгодженої рухової активності. Мозочок - головний керівний орган рухової системи, який здійснює координацію і контроль усіх видів рухів - від простих рухових актів, до складних форм поведінкової рухової активності.
Сіра речовина - кора - мозочка має три шари: поверхневий молекулярний шар, шар грушоподібних нейронів (клітини Пушкін'є) і нижній шар - зернистий. Крім кори в глибині мозочка серед білої речовини є скупчення нервових клітин, які утворюють його підкіркові парні ядра. Поверхня мозочка має велику кількість борозен, якими поділяється на окремі частини.
У шарах кори мозочка розміщені шість різних типів нейронів. Зерноподібні нейрони, кількість яких у людини становить розміщені у зернистомі шарі, а їхні аксони прямують до молекулярного шару, де розгалужуються Т-подібно, посилаючи в обох напрямках уздовж поверхні кори гілочки (паралельні нервові волокна) 1-2 мм завдовжки. Ці волокна проходять через ділянки розгалуження дендритів інших п'яти типів нейронів і утворюють на них синапси. У зернистому шарі розміщені також великі зірчасті нейрони (клітини Гольджі), дендрити яких поширюються на відносно невелику відстань, а аксони прямують до зерноподібних клітин.
Шар грушоподібних нейронів налічує у людини до 15 млн нейронів, дендрити яких розгалужуються у молекулярному шарі. Аксони грушоподібних нейронів спускаються до ядер мозочка, а невелика їх кількість закінчується на пристінкових ядрах. Аксони останніх клітин становлять єдиний еферентний шлях від мозочка.
Останні три типи нейронів - кошикові й зірчасті нейрони та клітини Лугаролежать у молекулярному шарі, причому напрямок аксонів клітин Лугаро невідомий. Аксони кошикових нейронів закінчуються на тілі, а зірчастих - на дендритах грушоподібних нейронів.
До кори мозочка входять два типи волокон. Висхідні нервові волокна (ліаноподібні) проходять крізь зернистий шар і закінчуються у молекулярному на дендритах грушоподібних нейронів, а дуже численні (у людини - до 50 млн)мохоподібні волокна закінчуються на зірчастих нейронах. Кожне мохоподібне волокно віддає велику кількість колатералей, завдяки чому воно іннервує кілька нейронів. До кожного нейрона кори мозочка підходять численні паралельні нервові волонка від зірчастих клітин, і тому через ці нейрони на будь-якій клітині кори мозочка конвертують сотні мохоподібних волокон.
Півкулі великого мозку через мозочок регулюють тонус скелетних м'язів і координують скорочення їх. Працями Л. А. Орбелі доведена участь мозочка в регуляції вегетативних функцій (діяльність серцево-судинної системи, дихання, травлення, терморегуляції). У новонароджених мозочок розвинений погано, найінтенсивніше росте в перший рік життя і до 15 років досягає розмірів дорослої людини.
Руйнування невеликих частин мозочка не має значного впливу на здійснення рефлекторний реакцій завдяки компенсації функції неушкодженими ділянками мозочка. При порушенні функції мозочка у людини виникає розлад координації рухів і м'язового тонусу. Типовим проявом уражень мозочка є ністагм (мимовільні, швидкі, ритмічні рухи очей), дрижання (тремор) і скандована мова, коли хворий нездатний координувати діяльність м'язів мовного апарату. Серед інших симптомів можуть спостерігатися: атонія - послаблення м'язового тонусу;астенія
- швидке стомлення і внаслідок цього зниження сили м'язових скорочень;адіадохокінез - уповільнена реакція під час зміни одного типу рухів на інший;дезеквілібрація - порушення рівноваги тіла; дистонія
- порушення тонусу м'язів; дисметрія - порушення амплітуди рухів;інтенційний тремор, або астазія, - проявляється коливанням і тремтінням рухів (м'язовий тремор). При ушкодженні мозочка виникають також запаморочення, деякі вегетативні порушення.
У тварин після травми мозочка з часом наступає досить ефективна компенсація його діяльності. Вона відбувається за рахунок функцій кори головного мозку, яка має двобічні зв'язки з мозочком. Однак поступове руйнування мозочка внаслідок будь-якого патологічного процесу супроводжується сильним головним болем і запамороченням, атаксією(порушення стояння і ходіння), що характеризується втратою узгодження між скороченням м'язів-синергістів, та м'язів-антагоністів. Хода хворого нагадує ходу п'яного, хворий широко розставляє ноги, ходить зигзагами, його "кидає" в обидва боки.
Водночас у людей з природженою вадою (відсутність мозочка) не спостерігається якихось істотних порушень рухових функцій. Це свідчить про те, що мозочок - не єдиний орган, який регулює рухову функцію організму, і коли його немає, це здійснюють структури довгастого, середнього та кінцевого мозку.
Між мостом і довгастим мозком спереду та мозочком ззаду розташований четвертий шлуночок. Дно четвертого шлуночка утворює ромбоподібна ямка, вкрита тонким шаром сірої речовини. Вона є задньою поверхнею моста й довгастого мозку. З боків ямка обмежована верхніми та нижніми ніжками мозочка.
По боках ромбоподібної ямки є два присередні підвищення. У верхній частині присереднього підвищення є лицевий горбок, утворений волокнами лицевого нерва, які оточують ядро відвідного нерва у вигляді коліна.
Знизу присереднє підвищення, поступово звужуючись, переходить у трикутник під'язикового нерва. Назовні від нього розташований трикутник блукаючого нерва.
У ділянці ромбоподібної ямки містяться ядра таких черепно-мозкових нервів: трійчастого, відвідного, лицевого, обох частин присінково-завиткового, язико-глоткового, блукаючого, додаткового, під'язикового.