ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ТРАНСПОРТЕ РЕСПИРАТОРНЫХ ГАЗОВ И УТИЛИЗАЦИИ КИСЛОРОДА В ОРГАНИЗМЕ
В физиологии труда и спорта приводится много данных о потреблении кислорода при мышечной деятельности, гораздо меньше об условиях транспорта респираторных газов.
Термин массоперенос, обозначающий процесс переноса массы вещества, нашел широкое распространение в литературе по физиологии дыхания. Сложный процесс массопереноса респираторных газов в организме, осуществляющийся в дыхательных путях и легких, системе кровообращения, в тканях и клетках и тесно связанный с утилизацией кислорода и образованием углекислого газа в условиях покоя, уже в значительной степени проанализирован в ряде монографий и других публикаций. Интересным представляется выявление особенностей этого процесса при мышечной деятельности, так как степень утилизации кислорода, образования углекислого газа и скорости массопереноса этих газов в организме оказываются наивысшими.
До недавнего времени основной характеристикой массопереноса респираторных газов было определение общего количества поглощенного организмом кислорода и выделенного углекислого газа. Этот подход, методика которого была предложена еще Холденом и Дугласом, представляет и в настоящее время большую ценность для физиологии дыхания. Однако, он характеризует лишь конечный результат процесса, не раскрывая его интимных механизмов.
Более детальную его характеристику дал методический подход, основы которого были заложены еще И.М. Сеченовым, К. Бором, А. Крогом и другими классиками физиологии дыхания, характеризующийся углубленным исследованием механизмов продвижения газов на отдельных участках их пути в организме. Этот подход получил дальнейшее развитие в современных условиях технического прогресса, на нем базируется изучение механизмов массопереноса респираторных газов.
Благодаря предложенной концепции о регулировании кислородных режимов организма (КРО), системному подходу к изучению дыхания, процесс массопереноса кислорода изучен у лиц разного возраста и пола при нормальном и измененном барометрическом давлении.
Сложное взаимодействие физиологических, физических и химических процессов, обусловливающих перенос газов внутри организма и между его внутренней и внешней средой, трудности методического характера ограничивают возможности использования только экспериментального подхода для непосредственного исследования этого процесса в живом организме. Зависимость процесса массопереноса респираторных газов в организме от состояния внешней среды, анатомических и физиологических условий, взаимозависимости нескольких физиологических систем, принимающих участие в регулировании этого процесса, пространственная неоднородность концентраций кислорода, углекислого газа и азота, неодинаковая скорость и разносторонняя направленность их движения, а также уровень функциональной активности органов, тканей или всего организма, обусловливают необходимость применения системного подхода, позволяющего вскрыть связи между изменениями в окружающей и внутренней среде, отдельными компонентами, управляющими воздействиями и параметрами процесса. Такой подход с использованием экспериментальных данных в настоящее время достаточно хорошо разработан (рис.1).
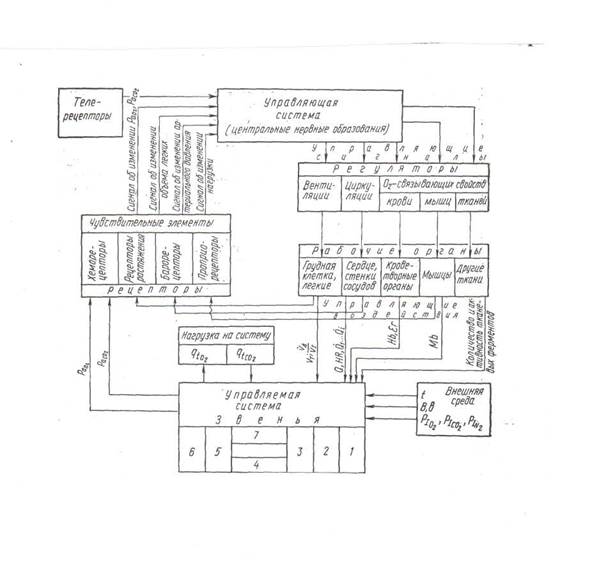
|
Рис. 1. Система дыхания.
Система дыхания рассматривается как состоящая из управляемой системы, в которой осуществляется процесс массопереноса респираторных газов, включающий несколько звеньев: кондуктивную и транзиторную зоны легких, респираторную зону легких, легочные капилляры, сосудистую систему с кровью, тканевые капилляры, клетки, венозную часть сосудистой системы; и управляющей системы: ЦНС и ее рабочими органами вентиляции, циркуляции; дыхательной функции крови, тканевыми механизмами и чувствительными элементами, передающими информацию об изменении параметров процесса в ЦНС.
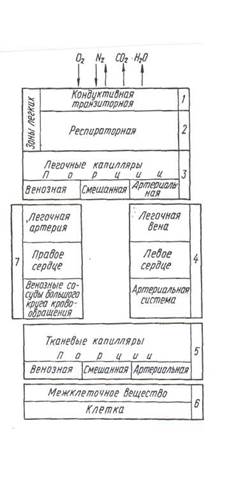
|
Рис. 2. Звенья управляемой системы дыхания
В первом звене управляемой системы (рис.2) – дыхательных путях, осуществляется движение кислорода с воздушным потоком к альвеолам, а углекислого газа, водяных паров, неиспользованного кислорода и инертных газов в обратном направлении – от альвеол к окружающей среде. Во втором звене – респираторной зоне легких, находящийся в альвеолах воздух вступает в контакт с кровью легочных капилляров. В третьем звене - происходит насыщение крови кислородом, в четвертом - благодаря сердечной деятельности кислород переносится от легких к тканям. Пятое звено - включает сеть капилляров в тканях и тканевой жидкости. Утилизация кислорода и образование углекислого газа и воды происходят в шестом звене – в клетках, и в седьмом – респираторные газы движутся по направлению к легким.
Благодаря контролю со стороны управляющей системы рабочие органы вырабатывают основные управляющие воздействия: периодическое изменение во времени объема легких, различные для разных тканей режимы кровоснабжения, поддержание необходимого количества эритроцитов, гемоглобина, миоглобина, дыхательных ферментов и коферментов. То есть, координация деятельности рабочих органов системы обеспечивается нейрогуморальной регуляцией функционирования внешнего дыхания, кровообращения, кроветворения и обменных процессов.
Одним из ведущих возмущающих воздействий на управление дыханием является кислородный запрос организма. Информацию об увеличении нагрузки, и связанной с этим необходимостью повышения потребления кислорода, центральные нервные образования получают как от проприорецепторов мышц и сухожилий, интерорецепторов различных органов, так и от хеморецепторов, реагирующих на изменения РО2 , РСО2 и рН.
В связи с тем, что массоперенос респираторных газов осуществляется многозвеньевой системой, необходимо понимать основные механизмы его регуляции в организме и особенности этого процесса при мышечной деятельности.
Скорость массопереноса газов в дыхательных путях зависит от целого ряда факторов. Еще в 1915 г. было показано, что у человека в состоянии покоя в дыхательных путях режим движения воздуха преимущественно ламинарный, при высоких скоростях потока значительное влияние оказывает турбулентность, делая режим движения воздуха смешанным. При увеличении вентиляции легких, когда скорость потока воздуха в легких увеличивается многократно (в процессе физических нагрузок, особенно интенсивных), турбулентность проявляется более резко. Так как массоперенос отдельных газов зависит, кроме скорости воздушного потока в легких, плотности воздуха, действия сил трения, инерционных сил, перепада барометрического давления, и от градиента концентраций (парциальных давлений газов), турбулентный режим движения воздуха значительно влияет на процесс диффузии газов и, таким образом, изменяет при мышечной деятельности условия массопереноса респираторных газов в легких.
Строение трахеобронхиального дерева, его геометрические размеры, изменяющиеся в каждом поколении, поперечное сечение воздухоносных путей, сопротивление воздушному потоку обусловливают последовательную или, как ее называют, послойную неравномерность транспорта и распределения газов между проксимальными и дистальными участками легких . Вследствие разного их растяжения, дихотомии дыхательных путей и связанных с этим неодинаковых изменений диффузионной поверхности и вентиляционно-перфузионных отношений в отдельных альвеолах при движении газов в различных зонах легкого возникает, кроме послойной, параллельная неравномерность распределения газов, что влияет на процесс их массопереноса.
С помощью математического моделирования выявлена временная характеристика конвективного и диффузионного массопереноса газов в каждой генерации бронхов в статике и динамике дыхательного цикла. Показано, что в одних отделах легких преобладает диффузионный массоперенос респираторных газов, в других - путем как диффузии, так и конвекции. Благодаря конвективному переносу, кислород премещается к 20-му поколению бронхов, дальнейшее его распространение осуществляется преимущественно путем диффузии. При малой скорости потока, когда в газовом транспорте преобладает диффузия, газовый состав в альвеолах cтановится более однородным.
Мышечная деятельность, в связи с изменением режима дыхания и необходимостью обеспечения переноса в единицу времени значительно большей, чем в покое, массы респираторных газов, изменяет соотношения между диффузионным и конвективным массопереносом газов. Укорочение дыхательного цикла при учащении дыхания уменьшает удельное значение диффузионного переноса газов при вентиляции альвеол.
Известно, что массоперенос кислорода из альвеол в кровь осуществляется через несколько неоднородных слоев: внутреннюю поверхность альвеол, покрытую сурфактантом, альвеолярный эпителий, базальные мембраны и капиллярный эндотелий. Кроме того, кислород и углекислый газ преодолевают диффузионное сопротивление плазмы крови и мембраны эритроцита. Скорость переноса газов через альвеолярно-капиллярную мембрану зависит от суммарной ее толщины и свойств составляющих ее компонентов, площади диффузионной поверхности, градиентов парциальных давлений газов, растворимости их в плазме крови, толщины и свойств мембраны эритроцита, способности гемоглобина и плазмы крови химически связывать газы и др. факторов. Процесс массопереноса респираторных газов на этом участке пути в организме осуществляется преимущественно путем диффузии.
В процессе мышечной деятельности, когда дыхательный объем увеличивается в 5 - 6 раз, в результате растяжения альвеол возрастает площадь диффузионной поверхности и уменьшается толщина альвеолярно-капиллярной мембраны, что в некоторой степени благоприятствует условиям диффузии респираторных газов. При этом, противоположно направленные потоки кислорода и углекислого газа взаимно влияют друг на друга, определяя скорость их массопереноса на участке между альвеолой и гемоглобином, зависящую от степени соединения газов с гемоглобином, скорости диссоциации угольной кислоты в эритроцитах и плазме, взаимодействия НСО3 с буферными системами плазмы и др. факторов.
Выявлено, что зависимость поступления кислорода в эритроцит от времени выражается экспонентой, и что при прохождении эритроцита по легочному капилляру в состоянии покоя он почти полностью насыщается кислородом приблизительно за 0,2 с. Время прохождения эритроцита по легочному капилляру при этом составляет около 0,7 с. При мышечной деятельности, когда скорость кровотока возрастает в 6-8 раз, времени контакта эритроцита с кислородом оказывается недостаточно для полного насыщения гемоглобина кислородом и, в связи с этим, условия для диффузии кислорода из альвеол в кровь в значительной степени меняются, в результате чего гемоглобин недонасыщается, снижая степень оксигенации крови легочных капилляров и артериальной крови (при интенсивных нагрузках на 15% и более), что ограничивает до некоторой степени доставку кислорода к тканям.
Несмотря на казалось бы достаточность времени для полной оксигенации гемоглобина в легких в покое, напряжение кислорода и углекислого газа в артериальной крови отличается от крови легочных капилляров из-за примеси венозной крови, поступающей из легочной артерии по шунтам и имеющей низкое РО2 и высокое РСО2, и неравномерности распределения газов в капиллярах различных участков легких. Объем легочного шунтирования может составлять в покое от 7 до 14% общего кровотока. Механизмы регуляции легочного шунта у здоровых людей следующие: анатомический шунт – сброс венозной крови по бронхиальным венам в легочные вены и по тебезиевым каналам в левое сердце; вентиляционно-перфузионная неравномерность или распределительные нарушения, т.е. перфузия гиповентилируемых либо гипоперфузия достаточно вентилируемых альвеол. При физической нагрузке значение шунтирования крови в легких усиливается вследствие значительного возрастания объемной скорости кровотока и, особенно, абсолютной величины анатомического шунта, в результате сброс венозной крови в артериальное русло может превышать в 5-7 раз величины покоя.
В способности гемоглобина обратимо присоединять кислород и углекислый газ важное значение имеют лиганды – вещества, связывающие атом железа гема с О2 и СО2: 2,3-дифосфоглицерат (ДФГ) и др.
Известно что синтез 2,3-ДФГ при нагрузках, превышающих 80% максимального потребления кислорода (МПК), затормаживается в результате накопления лактата.
Как известно, кривая диссоциации оксигемоглобина имеет S-образную форму, отражая способность гемоглобина насыщаться кислородом и отдавать его в довольно узкой области РО2. Количественным выражением сродства гемоглобина к кислороду являются, по предложению А. Крога, две точки: Р95, когда гемоглобин насыщен на 95%, и Р50 - на 50%. Установлено, что повышение Р50 на 3 мм рт.ст. способствует улучшению переноса кислорода к тканям (при тех же рН, РСО2 и температуре) приблизительно на 22%. Лишь при мышечной деятельности эти величины могут уменьшаться в значительной степени: в артериальной крови степень насыщения крови кислородом может снижаться до 78-80%, а в венозной – до 20% и менее.
Хорошо известна роль гемоглобина: насыщаясь кислородом, он становится более сильной кислотой, при этом осуществляет около 75% всего буферного действия крови, что имеет большое значение в регуляции кислотно-основного состояния крови (КОС), транспорте кислорода и углекислого газа. Особенно это важно при мышечной деятельности.
Повышение температуры и концентрации ионов водорода в крови при мышечной деятельности снижают сродство гемоглобина к кислороду, способствуют отдаче кислорода в тканях.
Ведущая роль в массопереносе респираторных газов от легких к тканям и обратно при мышечной деятельности принадлежит кровотоку, скорость которого многократно возрастает благодаря увеличению систолического объема (СО) и частоты сердечных сокращений (ЧСС). Немаловажное значение в обеспечении необходимой скорости доставки кислорода к работающим мышцам и выведения из них углекислого газа при мышечной деятельности имеет перераспределение кровотока, которое в первую очередь, связано с интенсивностью окислительных процессов в отдельных органах и тканях. Так, если в покое кислород в больших количествах поступает в мозг, сердце, печень и почки, то при физических нагрузках основная масса его доставляется к работающим мышцам. Наиболее информативным показателем этой связи может служить величина отношения между объемной скоростью кровотока и потреблением кислорода (гемодинамический эквивалент), которая при мышечной деятельности резко снижается.
Определенное значение для протекания процесса массопереноса респираторных газов в тканях имеют морфофункциональные особенности микроциркуляции крови. Установлена корреляция между функциональной активностью, уровнем метаболизма и капилляризацией ткани, которая зависит от степени двигательной активности. Важными факторами являются: число открытых капилляров (на 1 мм2), линейная скорость движения эритроцитов в капиллярах, их длина, время контакта эритроцита с окружающей тканью, градиент парциальных давлений газов вдоль капиллярной стенки.
На процесс массопереноса кислорода в тканях большое влияние оказывает миоглобин, т.к. присоединение кислорода к миоглобину поддерживает высокий градиент РО2 между цитоплазмой мышечной клетки и кровью. Это способствует ускорению диссоциации оксигемоглобина и поступлению молекулярного кислорода в цитоплазму. Высокое сродство миоглобина к кислороду, прямоугольно-гиперболическая форма кривой диссоциации оксимиоглобина обеспечивают непрерывность поступления кислорода в митохондрии при прерывистом потоке его подачи в клетки из крови. Кроме того, миоглобин оказывает катализирующее действие на скорость окислительных процессов в клетке.
Известно, что потребление кислорода в клетке определяется активностью целого комплекса кислородзависимых ферментов. Одним из самых серьезных последствий нарушения кислородного снабжения тканей является изменение условий для их работы. Несмотря на то, что перенос кислорода через клеточные мембраны осуществляется пассивно, на скорость его диффузии влияют состав и толщина самой мембраны и функциональное состояние клетки. Кривая зависимости скорости тканевого дыхания от концентрации кислорода является интегральным показателем, позволяющем оценить степень дисбаланса в работе кислородзависимых систем.
Скорость дыхания и скорость образования АТФ уменьшаются при РО2 в митохондриях -12 мм рт.ст. и меньше, а степень редукции цитохрома с (одного из главных ускорителей окислительно-восстановительных процессов в мышечных клетках) уменьшается уже при РО2 менее 40 мм рт.ст.
Процесс массопереноса респираторных газов в тканях зависит от их метаболических потребностей. На митохондриальном уровне он определяется величиной фосфатного потенциала, степенью редукции цитохрома с, наличием восстановленного и окисленного никотинамиддинуклеотида NAD – NAD/H.
В покое доставка кислорода к митохондриям избыточна по сравнению с их кислородным запросом, при мышечной деятельности, когда многократно увеличивается кислородный запрос и возникает несоответствие между доставкой кислорода и его потреблением, значительно снижается РО2 в мышцах.
Четкую количественную и качественную характеристику процесса массопереноса респираторных газов в организме дает анализ двух групп управляемых параметров: скорости движения газов на этапах их пути в организме – в легких, альвеолах, с артериальной и венозной кровью, потребления кислорода (выделения углекислого газа) тканями, а также парциальных давлений этих газов в альвеолах, крови и тканях . Их сочетания на каждом из этапов, строго управляемые и контролируемые в организме, названы кислородными режимами организма (КРО).
В организме в условиях устойчивого состояния устанавливается строгое соответствие поэтапной доставки кислорода кислородному запросу тканей. При этом количественный баланс (соответствие поэтапной доставки кислорода - кислородному запросу) при нормальном РО2 во вдыхаемом воздухе непосредственно определяет уровни парциального давления кислорода на разных участках пути в организме, зависит от отношения между скоростью доставки кислорода и скоростью его потребления и, в свою очередь, оказывает влияние на сам процесс.
Обеспечение тканей кислородом в различных условиях и при разных функциональных состояниях организма осуществляется неодинаковым уровнем функционирования органов внешнего дыхания, кровообращения. При изменении внешней среды (например, при снижении РО2 во вдыхаемом воздухе) и состояний организма (например, при мышечной деятельности) увеличивается расход кислорода. В таких случаях важно оценить: эффективность КРО, которая определяется по отношению скорости поэтапного продвижения кислорода к его потреблению; а также экономичность функционирования дыхания и кровообращения, которая определяется по отношению вентиляции легких и минутного объема крови к потреблению кислорода, между последним и частотой дыхания и сердечных сокращений (кислородные эффекты дыхательного и сердечного циклов).
Для полного представления о процессе массопереноса респираторных газов в организме необходим и анализ массопереноса углекислого газа, который можно охарактеризовать аналогично КРО.