Изменения возбудимости клетки во время её возбуждения
Билет 3
1. Рефлекс. Звенья рефлекторной дуги. Классификация рефлексов по центральному звену. Обратная связь, ее роль в рефлекторной деятельности на примере соматического рефлекса.
Рефлекс – стереотипная реакция организма в целом или его физиологических систем в ответ на раздражитель, реализуемая с помощью нервной системы.
Рефлекторная дуга – совокупность образований, необходимых для осуществления рефлекса.
5 звеньев: 1.рецептор 2.афферентное звено 3.центральное звено 4.эфферентное звено 5.эффектор.
По локализации центров – спинномозговые или спинальные, бульбарные ( с центральным звеном в продолговатом мозгу), мезенцефальные, диэнцефальные, мозжечковые, корковые.
Рефлекторная дуга замыкается в кольцо благодаря обратной афферентации – потоку импульсов от рецепторов эффектора обратно в рефлекторный центр. Функция обратной связи – сигнализация в центральную нервную систему о выполненном действии. Если оно выполнено недостаточно, нервный центр возбуждается – рефлекс продолжается. Также за счет обратной афферентации осуществляется контроль периферической деятельности центральной нервной системой.
Различают отрицательную и положительную обратные связи. Первая при выполнении определенной функции запускает механизм, угнетающий эту функцию. Положительная обратная связь заключается в дальнейшей стимуляции функции, которая уже выполняется или в угнетении функции, которая уже угнетена. Положительная обратная афферентация встречается редко, так как приводит биологическую систему в неустойчивое положение.
2.Мембранный потенциал покоя. Ионные механизмы, поддерживающие потенциал покоя клетки.
ММП - это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя.
1) Работа Na/K – насоса
2) Равновесный калиевый потенциал (= -97мВ). В клетку заходит меньше натрия, чем выходит калия, т.к проницаемость для калия в 10-15 раз больше, чем для натрия => обеспечение МПП
3) Органические анионы из-за больших размеров не могут выйти из клетки => обеспечение «-» заряда внутри
3. Нарисуйте схему саркомера, назовите сократительные и регуляторные белки, объясните значение ионов кальция в мышечном сокращении.
 актин
актин
тропонин-тропомиозиновый комплекс
миозин
Сократительные белки - актин и миозин.
Регуляторные белки –тропонин, тропомиозин.
Значение ионов кальция в мышечном сокращении: Ион кальция абсолютно необходим для нормального процесса сокращения. Соли кальция обладают положительным инотропным эффектом, а также влияют на тонус гладкой мускулатуры сосудов. В связи с тем, что ион кальция необходим для сокращения гладких мышц сосудов, он участвует в регуляции артериального давления путем действия на периферические сосуды.
4. Объясните разницу между первичным и вторичным торможением, назовите виды первичного торможения, нарисуйте схемы межнейронных контактов.
Торможение - местный нервный процесс, приводящий к угнетению или предупреждению возбуждения.
Первичное торможение (центральное) - результат возбуждения (активации) специальных тормозных нейронов.
·  Постсинаптическое торможение (прямое и возвратное)
Постсинаптическое торможение (прямое и возвратное)
· Пресинаптическое торможение
Вторичное торможение - осуществляется без участия специальных тормозных структур в тех самых нейронах в которых происходит возбуждение.
Типы межнейроннных контактов:
· аксоносоматические — между аксоном и клеткой ткани-мишени (н-р, как мы рисовали нервно-мышечный синапс)
· аксонодендритические — между аксоном и дендритом другого нейрона (самый обычный, как на картинке)
· аксоноаксональные — между данным аксоном и аксоном другого нейрона (рисунок пресинаптического торможения)
5. Дивергентные нейронные сети, их значение в деятельности ЦНС.
Дивергентные нейронные сети –сети, в которых контакты устанавливаются со все большим числом клеток последующих слоев иерархии. Обеспечивают интеграцию разных рефлекторных актов и общее состояние активности многочисленных нейронов разных отделов мозга.
Важную роль также играют так называемые дивергентные сети с одним входом. Командный нейрон, находящийся в основании такой сети может оказывать влияние сразу на множество нейронов, и поэтому сети с одним входом выступают согласующим элементом в сложном сочетании нейросетевых систем всех типов.
Билет 4
- Дайте сравнительную характеристику соматических и вегетативных рефлексов. Нарисуйте схему симпатической рефлекторной дуги.
| Соматическая нервная система | Вегетативная нервная система |
| Связь с органами однонейронная | Связь с органами двухнейронная: преганглионарный нейрон находится в центральной нервной системе, постганглионарный – в периферических ганглиях |
| Эффекторные нейроны находятся в центральной нервной системе | Эффекторные нейроны находятся на периферии (в ганглиях) |
| Эфферентные волокна идут только к скелетным мышцам | Эфферентные волокна иннервируют все органы, в том числе скелетные мышцы |
| Волокна выходят строго сегментарно, начиная от передних бугров четверохолмия среднего мозга и до конца спинного мозга | Волокна выходят из некоторых отделов центральной нервной системы: ядер среднего и продолговатого мозга, грудопоясничного отдела спинного мозга, крестцового отдела спинного мозга. Эти волокна называются преганглионарными; являются мякотными (миелинизированными), обеспечивают большую скорость проведения возбуждения |
| Волокна толстые, с большой скоростью проведения возбуждения (до 120 м/с); как правило, мякотные | Постганглионарные волокна тонкие, с малой скоростью проведения возбуждения (около 0,5 м/с), безмякотные (немиелинизированные) |
| Возбудимость высокая, быстро восстанавливается после возбуждения | Возбудимость низкая, медленно восстанавливается после возбуждения |
2 Потенциал действия. Нарисуйте график изменения мембранного потенциала и укажите ионные процессы при развитии ПД.Потенциал действия –быстрое колебание потенциала,сопровождающееся ,как правило,перезарядкой мембраны

3.Нарисуйте схему саркомера, назовите сократительные и регуляторные белки, объясните значение АТФ в мышечном сокращении.

Основными сократительными белками саркомера являются миозин, актин и тайтин. Молекулы актина образуют тонкие (актиновые) нити, в состав которого входят такжерегуляторные белки тропомиозин и тропонин. Толстые нити скелетных мышц кроме миозина содержат также другие белки, такие как тайтин, С-белок, Х-белок, Н-белок
Универсальным источником энергии в живом организме является молекула АТФ. Которая превращаясь в «бесполезную» АДФ дает нам самую удобную для потребления энергию.
АТФ + H2O = АДФ и кислота + энергия. «Бесполезная» молекула АДФ способна снова превратиться в «годную» АТФ если есть достаточное количество креатинфосфата
АДФ + креатинфосфат = АТФ + креатин.
Важная особенность заключается в том, что на востонавление запасов креатинфосфата нужно несколько минут и то, что эта реакция может осуществляться только после прекращения работы. Если бы креатинфосфат мог восстанавливаться во время работы, мы бы могли долго работать тяжелыми весами в очень большом количестве повторений.
Билет 5
5 билет
1 вопрос.
| Нервная | Гуморальная |
| Распространение нервного импульса по нервным волокнам | Распространение химического вещества с током крови |
| Нервный импульс распространяется с очень высокой скоростью по нервным волокнам | Импульс в виде химического вещества распространяется со скоростью кровотока |
| Эффект строго ограничен (локализован) изменением функционирования того органа или ткани, который получил «команду» в виде нервного импульса | Эффект реализуется широко (генерализован) и проявляется во всех органах, где есть клетки-мишени для данного гормона |
| Длительность действия ограничена быстрым специфическим ответом клетки – сокращение мышцы, выделение секрета. | Длительность действия увеличивается, во-первых, временем циркуляции гормона в крови до его разрушения, во-вторых, длительными изменениями функционирования клеток, которые обусловлены изменением метаболизма и даже структуры клеток. |
 Цифрой 1 на рисунке 3 обозначены осморецепторы печени, которые возбуждаются при увеличении осмотической концентрации внутренней среды. Информация от этих рецепторов поступает к супраоптическим ядрам гипоталамуса. Нейроны этих ядер способны синтезировать гормон, который из за своей способности снижать выделение мочи получил название антидиуретический (АДГ). В нашем примере именно гипоталамические ядра с эндокринной функцией и бкдкт эффектором номер1 (цифра 2 на рисунке). Этот гормон, поступивший по аксонам гипоталамических нейронов в заднюю долю гипофиза выделяется в кровь и действует на собирательные трубочки нефронов - это эффектор второго порядка (3) - таким образом, что они становятся проницаемы для воды. Вода задерживается в организме, приводя к восстановлению осмотическую концентрацию внутренней среды. АДГ оказывает свое влияние и на кровеносные сосуды – это еще один эффектор второго порядка, и на центральную нервную систему - формирует чувство жажды и соответствующее поведение. В гипоталамусе осуществляется наиболее важная в физиологическом плане связь нервной и эндокринной систем - происходит трансформация нервного импульса в специфический эндокринный процесс. В свою очередь, выделяющиеся гормоны оказывают влияние на чувствительность нервных клеток, скорость проведения нервного импульса в ЦНС.
Цифрой 1 на рисунке 3 обозначены осморецепторы печени, которые возбуждаются при увеличении осмотической концентрации внутренней среды. Информация от этих рецепторов поступает к супраоптическим ядрам гипоталамуса. Нейроны этих ядер способны синтезировать гормон, который из за своей способности снижать выделение мочи получил название антидиуретический (АДГ). В нашем примере именно гипоталамические ядра с эндокринной функцией и бкдкт эффектором номер1 (цифра 2 на рисунке). Этот гормон, поступивший по аксонам гипоталамических нейронов в заднюю долю гипофиза выделяется в кровь и действует на собирательные трубочки нефронов - это эффектор второго порядка (3) - таким образом, что они становятся проницаемы для воды. Вода задерживается в организме, приводя к восстановлению осмотическую концентрацию внутренней среды. АДГ оказывает свое влияние и на кровеносные сосуды – это еще один эффектор второго порядка, и на центральную нервную систему - формирует чувство жажды и соответствующее поведение. В гипоталамусе осуществляется наиболее важная в физиологическом плане связь нервной и эндокринной систем - происходит трансформация нервного импульса в специфический эндокринный процесс. В свою очередь, выделяющиеся гормоны оказывают влияние на чувствительность нервных клеток, скорость проведения нервного импульса в ЦНС.
2 вопрос
При действии раздражителя подпороговой силы возникает неполная деполяризация - ЛОКАЛЬНЫЙ ОТВЕТ (ЛО). Неполная, или частичная деполяризация – это такое изменение заряда мембраны, которое не достигает критического уровня деполяризации (КУД).
1.  МПП - электроотрицательность мембраны клетки в покое, обеспечивающая способность к возбуждению - возбудимость. На рисунке МПП = -90 мв.
МПП - электроотрицательность мембраны клетки в покое, обеспечивающая способность к возбуждению - возбудимость. На рисунке МПП = -90 мв.
- КУД - критический уровень деполяризации (или порога генерации мембранного потенциала действия) - это такая величина мембранного потенциала, при достижении которой открываются быстрые, потенциал зависимые натриевые каналы и происходит перезарядка мембраны за счет поступления в клетку положительных ионов натрия. Чем выше электроотрицательность мембраны, тем труднее деполяризовать ее до КУД, тем менее возбудима такая клетка.
- Точка реверса потенциала (овершут) - такая величинаположительногомембранного потенциала, при которой положительно заряженные ионы уже не проникают в клетку - кратковременный равновесный натриевый потенциал. На рисунке + 30 мв. Суммарное изменение потенциала мембраны от –90 до +30 составит для данной клетки 120 мВ, эта величина и является потенциалом действия. Если этот потенциал возник в нейроне, он будет распространяться по нервному волокну, если в мышечных клетках – будет распространяться по мембране мышечного волокна и приведет к сокращению, в железистых к секреции – к действию клетки. Это и есть специфический ответ клетки на действие раздражителя, возбуждение.
3 ВОПРОС
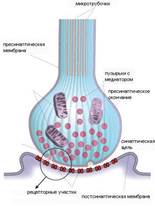
Передача нервного импульса с нервного волокна на другую клетку осуществляется посредством СИНАПСОВ. Синапс - специализированная зона контактамежду аксоном и другим нейроном, мышечной или секреторной клеткой, обеспечивающая передачу нервного импульса с нервного волокна на эффекторную клетку
Механизм нервно-мышечной передачи
Синапсы представляют собой коммуникационные структуры, которые формируются окончанием нервного волокна и прилегающей к нему мембраной мышечного волокна (пресинаптической нервной и постсинаптической мышечной мембранами)
Когда нервный импульс достигает окончания аксона, на деполяризованной пресинаптической мембране открываются потенциалзависимые Са2+ каналы. Вход Са2+ в аксональное расширение (пресинаптическую мембрану) способствует высвобождению химических нейромедиаторов, находящихся в виде везикул (пузырьков) из окончания аксона. Медиаторы (в нервно-мышечном синапсе это всегда ацетилхолин) синтезируются в соме нервной клетки и путем аксонального транспорта транспортируются к окончанию аксона, где и выполняют свою роль. Медиатор диффундирует через синаптическую щель и связывается со специфическими рецепторами на постсинаптической мембране. Так как медиатором в нервно-мышечном синапсе является ацетилхолин, то рецепторы постсинаптической мембраны называют холинорецепторами. В результате этого процесса на постсинаптической мембране открываются хемочувствительныеNа+-каналы, возникает деполяризация, величина которой различна, и зависит от количества выделенного медиатора. Чаще всего возникает локальный процесс, который называют потенциалом концевой пластинки (ПКП). При повышении частоты стимуляции нервного волокна, усиливается деполяризация пресинаптической мембраны, а, следовательно, возрастает количество выделяемого медиатора и число активированных хемочувствительныхNа+каналов на постсинаптической мембране. Таким образом, возникают ПКП, которые по амплитуде деполяризации суммируются до порогового уровня, после чего, на мембране мышечного волокна, окружающей синапс, возникает ПД, который обладает способностью к распространению вдоль мембраны мышечного волокна. Чувствительность постсинаптической мембраны регулируется активностью фермента – ацетилхолинэстеразы (АЦХ-Э), который гидролизует медиатор АЦХ на составные компоненты (ацетил и холин) и возвращает назад – в пресинаптическую бляшку для ресинтеза. Без удаления медиатора на постсинаптической мембране развивается длительная деполяризация, которая ведет к нарушению проведения возбуждения в синапсе – синаптической депрессии. Таким образом, синаптическая связь обеспечивает одностороннее проведение возбуждения с нерва на мышцу, однако на все эти процессы расходуется время (синаптичекая задержка), что приводит к низкой лабильности синапса по сравнению с нервным волокном.
Таким образом, нервно-мышечный синапс является «выгодным» местом, куда можно воздействовать фармакологическими препаратами, изменяя чувствительность рецептора, активность фермента. Эти явления будут часто встречаться в практике врача: например, при отравлении токсином ботулизма – блокируется высвобождение медиатора АЦХ (разглаживание морщин в косметической медицине), блокада холиноререпторов (курареподобными препаратами, бунгаротоксином) нарушает открытие Nа+ каналов на постсинаптической мембране. Фосфоорганические соединения (множество инсектицидов) нарушает эффективность АЦХ-Э и вызывает длительную деполяризацию постсинаптической мембраны. В клинике используют специфические блокаторы нервно-мышечного проведения: блокада холинорецепторов курареподобными препаратами, сукцинилхолином и другими конкурентными ингибиторами, вытесняющими АЦХ с холинорецептора.При заболевании миастении из-за дефицита холинорецепторов на постсинаптической мембране (из-за их аутолитического разрушения) возникает прогрессирующая мышечной слабость, вплоть до полной остановки мышечных сокращений (остановка дыхания). В этом случае используют блокаторы АЦХ-Э, что приводит к увеличению длительности связывания медиатора с меньшим количеством холинорецепторов и несколько увеличивает амплитуду деполяризации постсинаптической мембраны.
6 билет
1. Классификация рефлексов по рецепторному звену. Роль рецепторов в осуществлении рефлекса. Понятие рецептивного поля и рефлексогенной зоны.
2. Объясните, каким образом и почему изменяется возбудимость клетки в соответствии с фазами потенциала действия.
1. Классификация по рецепторному звену.
· Интероцептивные: информация, возбуждающая рецептор и, запускающая рефлекс, получена с рецепторов внутренних органов;
· Экстероцептивные: информация, возбуждающая рецептор и, запускающая рефлекс, получена из внешней среды с помощью сенсорных систем;
· Проприоцептивные: рефлексы, запускаемые с рецепторов мышц, сухожилий и суставов.
· по центральному звену выделяют: центральные (истинные) – главное звено находится в ЦНС и периферические – центральное звено находится за пределами ЦНС.
Центральные, в свою очередь делятся на спинальные и церебральные. Спинальные рефлексы делят на цервикальные, торакальные, люмбальные и сакральные. Церебральные рефлексы делят на мозжечковые, рефлексы больших полушарий и рефлексы ствола мозга. Рефлексы ствола мозга делят на бульбарные, диэнцефальные и мезенцефальные.
Структурной основой осуществления рефлекса является так называемая рефлекторная дуга. В ее состав входят: рецепторы, воспринимающие воздействующие на организм раздражители; афферентные нейроны, которые своими отростками связывают рецепторы с центрами нервной системы и обеспечивают центростремительное проведение возбуждения; структуры спинного и головного мозга, куда поступает афферентное возбуждение и где происходит его анаэфферентные нейроны, являющиеся конечным пунктом выхода из ЦНС возбуждения, распространяющегося центробежно по нервным волокнам на периферию к органам и тканям; эффекторы, или исполнительные органы (скелетные мышцы, железы, внутренние органы), реализующие рефлекс. Конкретные рефлекторные реакции могут включать различное количество рецепторов, афферентных и эфферентных нейронов и сложные процессы взаимодействия возбуждений в нервных центрах. Области тела, в пределах которых расположены специализированные рецепторы, раздражение которых вызывает строго определенные рефлексы, получили название рецептивных (рецепторных) полей, или рефлексогенных зон. Например, раздражение слизистой оболочки носа вызывает рефлекс чихания, а слизистой оболочки глазного яблока – мигательный рефлекс. Рефлексогенные зоны формируются в соответствии с врожденной программой развития и созревания нервной системы индивидуума.
Афферентный путь проведения возбуждения от рецепторов в ЦНС может включать от одной до нескольких афферентных нервных клеток. Первая нервная клетка, непосредственно связанная с рецептором, называется рецепторной, последующие – часто называют сенсорными, или чувствительными. Они могут располагаться на различных уровнях ЦНС, начиная от спинного мозга и кончая афферентными зонами коры больших полушарий. Афферентные нервные волокна, являющиеся отростками рецепторных нейронов, проводят возбуждение от рецепторов с различной скоростью.
Механизмы формирования и осуществления рефлекса связаны со структурнофункциональными особенностями всех звеньев рефлекторной дуги. Интервал времени от момента воздействия раздражителя на рецепторы до начала изменения функционального состояния эффектора обозначается как латентный (скрытый) период, или время рефлекса. Например, у человека время коленного рефлекса составляет 0,0196-0,0238 с, время мигательного рефлекса – 0,05-0,2 с, время рефлекса расширения кожных сосудов достигает 20 с.
2. Все клетки организма имеют электрический заряд, обеспечиваемый неодинаковой концентрацией анионов и катионов вне и внутри клетки. При действии раздражителя на клетку возбудимой ткани изменяется проницаемость ее мембраны, вследствие чего ионы быстро перемещаются согласно электрохимическому градиенту (совокупность концентрационного и электрического градиентов), - это и есть процесс возбуждения. Его основой является потенциал покоя.
Изменения возбудимости клетки во время её возбуждения
Возбудимость клетки во время ее возбуждения быстро и сильно изменяется.Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и, так же как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов. Схематично эти изменения представлены на рисунке.

Рисунок .Фазовые изменения возбудимости клетки (b) во время ПД (а).
1,4 - возбудимость повышена; 2 - абсолютная рефрактерная фаза;3 - относительная рефрактерная фаза
Билет 6.3: Возбудимость мышцы во время одиночного мышечного сокращения изменяется в соответствии с фазами потенциала действия - мышца невозбудима в период абсолютной рефрактерности, соответствующий пику ПД. По времени этот период приблизительно совпадает с латентным периодом в мышечном сокращении. Следовательно, во время одиночного мышечного сокращения возбудимость скелетной мышцы остается на нормальном уровне, а значит, мышца может ответить на раздражение в течение всего времени одиночного сокращения.
 Рис.Временная характеристика процессов возбуждения и сокращения
Рис.Временная характеристика процессов возбуждения и сокращения
АМПЛИТУДА одиночного сокращения всей скелетной мышцы зависит от количества сократившихся двигательных единиц. Возбудимость отдельных групп волокон, составляющих целую мышцу, различна и поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых волокон. Амплитуда такого сокращения незначительна. Амплитуда сокращения становится максимальной тогда, когда в процесс возбуждения вовлекаются все мышечные волокна, когда превышен порог раздражения всех двигательных единиц, составляющих мыщцу. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается при дальнейшем нарастании силы раздражающего тока.
Если интервал между раздражениями превышает длительность одиночного сокращения, мышца успевает полностью расслабиться. Однако если увеличить частоту импульсов тока, то каждый последующий импульс совпадает с фазой расслабления мышцы. Поскольку мышца в этот период имеет нормальную возбудимость, то амплитуда сокращений будет суммироваться и возникнет ЗУБЧАТЫЙ ТЕТАНУС - слитное сокращение мышцы, когда она не расслабляется до исходной величины. При дальнейшем увеличении частоты раздражения каждый последующий импульс тока действует на мышцу в тот период, когда она находится в фазе укорочения. Возникает ГЛАДКИЙ ТЕТАНУС - длительное укорочение, не прерываемое расслаблением.
 Зубчатый тетанус. Сокращения мышцы без расслабления, раздражение совпадает с фазой расслабления. Мышца не успевает расслабиться полностью перед следующим импульсом раздражения..
Зубчатый тетанус. Сокращения мышцы без расслабления, раздражение совпадает с фазой расслабления. Мышца не успевает расслабиться полностью перед следующим импульсом раздражения..
 Гладкий тетанус. Раздражение с такой частотой, при которой действие раздражителя совпадает с фазой укорочения. Мышца не успевает даже начать расслабляться перед следующим импульсом раздражения.
Гладкий тетанус. Раздражение с такой частотой, при которой действие раздражителя совпадает с фазой укорочения. Мышца не успевает даже начать расслабляться перед следующим импульсом раздражения.
Билет 6.4: Виды первичного торможения:
Нервные импульсы, возникающие при возбуждении тормозных нейронов, вызывают гиперполяризацию постсинаптической мембраны и тормозной постсинаптический потенциал (ТПСП), это наиболее распространенный тип торможения в нервной системе, который называется ПОСТСИНАПТИЧЕСКИМ ТОРМОЖЕНИЕМ. Постсинаптическое торможение является первичным торможением – самостоятельный процесс, вызванный возбуждением тормозных нейронов.
Существует еще один вариант первичного торможения в ЦНС: ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ. Такое торможение возникает в том случае, когда нервный импульс или просто не достигает синаптического окончания аксона, или снижается его частота. Такое торможение осуществляется благодаря аксоаксональным синапсам, образованным терминалями аксонов тормозных интернейронов и аксонами возбуждающих нейронов (рис.27, справа). В этих синапсах торможение определяется действием ГАМК, открывающей хлорные каналы на пресинаптических мембранах (мембрана возбуждающего нейрона). Из - за высокого МПП мембраны аксона (действие электрического, а не концентрационного градиента) ионы хлора выходят через мембрану наружу. Выход ионов вызывает незначительную деполяризацию постсинаптической мембраны, являющейся в данном случае участком мембраны аксона другого нейрона. Деполяризация мембраны аксона ведет к инактивации натриевой проводимости. Возникает блокирование проведения нервного импульса, что уменьшает частоту, или полностью угнетает возбуждающий потенциал, идущий к клетке по возбуждающим нервным окончаниям.
| Постсинаптическое торможение | Пресинаптическое торможение |

| 
|
| Возбудимость нейрона 1 снижена в результате гиперполяризации мембраны, вызванной действием тормозного нейрона, поэтому два возбуждающих нейрона не могут вызвать ПД. | Возбудимость нейрона 1 не изменена, но возбуждающий импульс от одного из возбуждающих нейронов не доходит, а один возбуждающий не может вызвать ПД. |
Первичное торможение
Вторичного торможения:
Состояния нейрона
На рисунке представлены состояния нейрона: поляризован - обладает мембранным потенциалом покоя, гиперполяризован и деполяризован – потенциал действия. Пока не расшифровывая аббревиатуры «ТПСП», вспомним, что гиперполяризация – это следовой процесс, возникающий при восстановлении МПП после возбуждения. Гиперполяризованный нейрон менее возбудим, потому, что потенциал его мембраны «уходит» от критического уровня деполяризации. Таким образом, снижение возбудимости нейрона может быть результатом длительного возбуждения. Кроме того, возможно простое утомление нейронов, и особенно, синапсов – расходуются запасы АТФ, необходимые для восстановления МПП, запасы медиатора. Следовательно, торможение может быть результатом длительной работы. Такой вариант торможения распространен в ЦНС и получил название вторичного торможения. Один из видов вторичного торможения получил название ПЕССИМАЛЬНОЕ ТОРМОЖЕНИЕ (ТОРМОЖЕНИЕ ВВЕДЕНСКОГО). Такое торможение развивается в результате действия раздражителей с очень высокой частотой. В результате клетка не успевает восстановить свой МПП, остается деполяризованной и, следовательно, невозбудимой 
Межнейронные контакты: Нейроны за счет ветвления коллатералей аксона и установления множества синаптических контактов между нервными клетками объединяются в цепочки и сети. Цепочки могут быть простые и локальные. Нейронные сети называются иерархическими потому, что передача нервного импульса происходит от уровня к уровню. Сети бывают конвергентные и дивергентные.
Простые нейронные цепочки – это путь распространения импульса от нейрона к нейрону. В локальной цепочке, которая еще называется кольцевая цепь Лоренто де Но, импульс может циркулировать по замкнутому кругу достаточно долго (рис.28). Возвращение возбуждения к «первому» нейрону локальной сети получило название реверберациивозбуждения. Функциональное значение реверберации импульса заключается в пролонгировании распространения ПД – есть нервные центры, например, гиппокамп, где реверберация может продолжаться несколько дней. Такое самоподдерживающееся распространение ПД играет большую роль в формировании следа памяти.
