IV. Гипотеза А.М. Оловникова
КАРАГАНДИНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
Кафедра молекулярной биологии и медицинской генетики
СРС
На тему: «Теломеры. Теломеразная активность.»
Выполнила: ст. группы 1-032 ОМ
Таскынбай А.Е.
Проверила: Авдиенко О.В.
Караганда 2016г.
Цель:Изучение теломеразных последовательностей ДНК, функций теломер, механизма действия теломераз.
Задачи:
1) Изучить структуру теломер.
2) Функции теломер.
3) Методы определения активности теломеразы
Содержание:
I. Введение……………………………………………………..4 стр.
II. Теломера: строение, функции……………………………...5 стр.
III. «Концевая недорепликация ДНК» ………………………..6 стр.
IV. Гипотеза А.М. Оловникова ………………………………..6 стр.
V. Теломераза. Действие теломеразы………………………6-7 стр.
VI. Теломеразная активность клеток ……………………….7-8 стр.
VII. Заключение………………………………………………….9 стр.
VIII. Список используемой литературы ………………………10 стр.
I. Введение
На концах линейных хромосом эукариот находятся специализированные ДНК-белковые структуры – теломеры (Оловников А.). У большинства организмов теломерная ДНК представлена многочисленными короткими повторами. Их синтез осуществляется необычным РНК-содержащим ферментом теломеразой. В презентации обсуждаются связь длины теломерных районов хромосом и активности теломеразы с процессами старения и злокачественного роста. Изучение тонкой структуры теломер и механизма действия теломераз находится еще только в начальной стадии. Однако они привлекают к себе огромный интерес исследователей, работающих в самых разных областях биологии и медицины, и здесь уже в ближайшее время можно ждать новых интересных открытий.
Существование структур на концах хромосом было открыто в 1938 году классиками генетики, лауреатами Нобелевской премии Барбарой Мак-Клинток и Германом Мёллером.Независимо друг от друга они обнаружили, что фрагментация хромосом (под действием рентгеновского облучения) и появление у них дополнительных концов ведут к хромосомным перестройкам и деградации хромосом.
В сохранности оставались лишь области хромосом, прилегающие к их естественным концам. Лишенные концевых теломер, хромосомы начинают сливаться с большой частотой, что ведет к тяжелым генетическим аномалиям. Следовательно, заключили они, естественные концы линейных хромосом защищены специальными структурами.
В 1932 Г. Мёллер предложил называть их теломерами (от греч. телос - конец и мерос - часть).
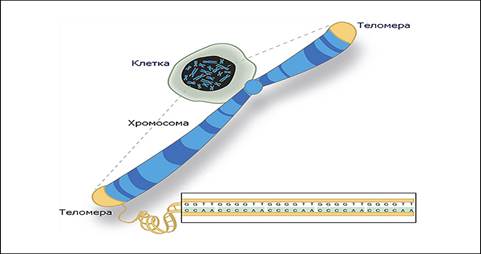
Хромосома имеет две теломеры. У человека теломеры содержат единственный повтор GGGTTA.
II. Теломера: строение, функции
В клетках человека теломеры обычно представлены одноцепочечной ДНК и состоят из несколько тысяч повторяющихся единиц последовательности ТТАГГГ. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые
G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом.
Теломерная ДНК имеет определенный состав, а для поддержания ее длины, как правило, используется специальный фермент — теломераза.
Нуклеотидная структура теломерных последовательностей ДНК нам уже известна.
Более высокие уровни организации образуются за счет специфических белков.
Благодаря этим белкам, теломеры имеют плотную упаковку, т. е. относятся к фракции гетерохроматина.
С помощью теломерных белков теломеры крепятся к компонентам ядерного матрикса, в т. ч., возможно, к ядерной ламине.
Полагают также, что теломерная ДНК образует несколько петель (в виде «лепестков ромашки»), фиксированных на матриксе; и по мере укорочения теломер число
«лепестков» постепенно уменьшается.
Функции теломер:
• Механические: теломеры участвуют в фиксации хромосом к ядерному матриксу. Это важно для правильной ориентации хромосом в ядре, и данное обстоятельство особенно проявляется в мейозе
• Стабилизационные: В присутствии теломеразы к местам разрыва присоединяется теломерная ДНК. Это стабилизирует хромосомные фрагменты и позволяет им функционировать.
• Отсчет количества клеточных делений: отсчитывает количество делений клетки после исчезновения теломеразной активности
III. «Концевая недорепликация ДНК»
ДНК-полимеразы, синтезируя дочернюю цепь ДНК, прочитывают родительскую цепь в направлении от ее 3'-конца к 5'-концу. Соответственно дочерняя цепь синтезируется в направлении 5' 3'. В противоположном направлении синтез цепи ДНК фермент катализировать не может. ДНК-полимераза начинает синтез только со специального РНК-праймера - короткой РНК-затравки, комплементарной ДНК. После окончания синтеза ДНК РНК-праймеры удаляются, а пропуски в одной из дочерних цепей ДНК заполняются ДНК-полимеразой. Однако на 3'-конце ДНК такой пропуск заполнен быть не может, и поэтому 3'-концевые участки ДНК остаются однотяжевыми, а их 5'-концевые участки - недореплицированными. Отсюда ясно, что каждый раунд репликации хромосом будет приводить к их укорочению. Понятно, что прежде всего должна сокращаться длина теломерной ДНК.
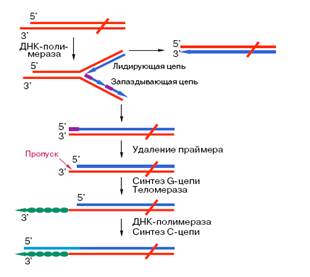
IV. Гипотеза А.М. Оловникова
Первым на проблему "концевой недорепликации ДНК" обратил внимание А.М. Оловников в 1971 году.
Он высказал гипотезу о том, что потеря концевых последовательностей ДНК вследствие их недорепликации ведет к старению клетки.
Иными словами, предполагалось, что процесс укорочения теломер и есть тот часовой механизм, который определяет репликативный потенциал "смертной" клетки, и когда длина теломер становится угрожающе короткой, этот механизм предотвращает дальнейшее деление клетки.
А.М. Оловников предположил также, что в нестареющих клетках (а к ним кроме раковых относятся зародышевые, стволовые и другие генеративные клетки) должна существовать специализированная ферментативная система, которая контролирует и поддерживает длину теломерной ДНК.