Цепной миотатический рефлекс
Глава 5. Клинико-патогенетическое значение цепных миотатических синкинезий
Особенности функциональной анатомии поперечнополосатых и гладких мышц
Скелетные (поперечнополосатые) мышцы состоят из множества функциональных единиц – мышечных волокон (клеток). Они имеют цилиндрическую форму и тесно прилегают друг к другу. Диаметр клеток поперечнополосатых мышц составляет от 0,005 до 0,1 мм, а длина может достигать нескольких сантиметров. Пучки мышечных волокон окружены соединительнотканными прослойками. На конце мышц коллагеновые волокна соединительной ткани образует сухожилия, которые служат для прикрепления мышц к разным частям скелета.
Общее число соединительной ткани в скелетной мышце довольно значительно. В зависимости от типа мышцы содержание коллагена в ней колеблется от 30% до 3% (рис. 37).
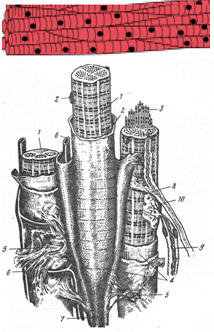
Рис. 37. Схема строения поперечно-полосатой мышечной ткани.
1 - мышечные волокна; 2 - ядра; 3 - миофибриллы; 4 - сарколемма; 5 - эндомизий; 6 - кровеносные капилляры; 7 - сухожильная нить; 8 - вегетативное нервное волокно; 9 - соматическое (двигательное) нервное волокно; 10 - аксоно-мышечный синапс [78]
Соединительнотканные прослойки мышцы обильно иннервируются рецепторами (колбами Краузе, тельцами Фатера-Пачини, ноцицепторами). Они располагаются и в фасциальных нервных стволах, и в жировых дольках.
Поперечнополосатые мышцы вместе со связками и периартикулярными тканями составляют около половины массы человеческого тела.
Мышечная клетка (волокно) одета прозрачной оболочкой – сарколеммой. Последняя по структуре подобна мембране нервной клетки и играет важную роль в возникновении и проведении возбуждения.
Цитоплазма мышечной клетки (саркоплазма) состоит из саркоплазматического матрикса – однородной среды, в которую погружены миофибриллы – сократимая субстанция неживой природы. Одно волокно содержит до 1000 фибрилл [68].
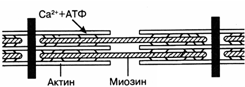
Рис. 24.Схема сокращения актин-миозинового комплекса
Каждая из миофибрилл состоит из саркомеров, образованных белковыми нитями двух типов - актином и миозином. При сокращении мышцы укорачиваются лишь волокна миозина. Они разбухают в поперечном направлении благодаря поглощению воды. При растяжении мышцы миозиновые столбики удлиняются. Сокращение актин-миозинового комплекса возможно только при присоединении ионов C2+ к особому регуляторному белку тропонину, локализованному на миозиновых нитях. Кроме того, для этого необходимо расщепление одной молекулы АТФ (рис. 38).
Мышечные волокна, осуществляющие непосредственное сокращение мышцы, называются экстрафузальными. Различают ещё и специализированные – интрафузальные - мышечные волокна, которые непосредственно в сокращении мышцы не участвуют. Они входят в состав сенсорных образований мышцы или её проприорецепторов – мышечных веретён.
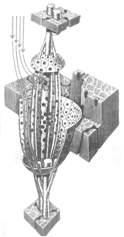
Рис. 25. Модель типичного мышечного веретена, сокращение саркомера [255]
В каждой мышце насчитывается от 90 до 100 мышечных веретён, которые расположены параллельно экстрафузальным мышечным волокнам и являются своего рода датчиками растяжения мышцы. Типичное мышечное веретено состоит из нескольких интрафузальных волокон, заключённых внутри толстой соединительнотканной капсулы (рис. 39).
Экстрафузальные мышечные волокна получают двигательную иннервацию от альфа-, а интрафузальные – от гамма-мотонейронов переднего рога спинного мозга. Кроме того, интрафузальные волокна веретён снабжены ещё и специализированными чувствительными (афферентными) нервными окончаниями.
Суть рефлекса на растяжение мышцы (миотатического рефлекса) заключается в следующем: любое увеличение длины мышцы сопровождается растяжением веретён. При этом происходит возбуждение афферентов. Возникающий импульс приходит к альфа-мотонейронам, вызывая сокращение всей мышцы за счёт её экстрафузальных волокон. Степень сокращения определяется величиной предшествующего растяжения.
Вследствие действия внешних факторов (инерция, гравитация) миотатический рефлекс минимальной степени выраженности наблюдается постоянно во всех скелетных мышцах, даже в условиях полного покоя и играет важную роль в поддержании и регуляции мышечного тонуса [60].
Помимо сократимости, важными биомеханическими свойствами скелетных мышц являются упругость и растяжимость.
Эластические структуры мышцы аккумулируют энергию упругих деформаций и частично возвращают её во время движения. Например, в ахилловом сухожилии может накапливаться потенциальной энергии упругой деформации больше, чем в икроножной мышце. Однако длительно растянутая и затем отпущенная мышца возвращается к своей исходной длине не полностью, а остается некоторое время более или менее удлиненной. Все эти явления связаны с вязкоупругими свойствами соединительнотканных элементов мышцы [33].
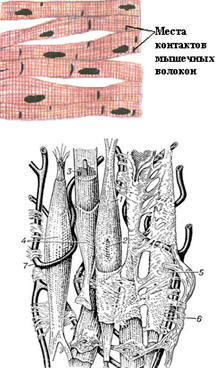
Рис. 26. Схема строения гладкой мышечной ткани.
1 - гладкая мышечная клетка; 2 - её ядро; 3 - миофибриллы; 4 - сарколемма; 5 - соединительная ткань; 6 - нерв; 7 - кровеносный капилляр [78]
Гладкая мышечная ткань, так же, как и поперечнополосатые мышцы, имеет клеточное строение. Её клетки толщиной 2-10 мкм имеют веретенообразную форму и небольшую, по сравнению со скелетными волокнами, длину – 50 - 400 мкм.
Гладкие мышцы контактируют друг с другом отростками так, что мембраны двух клеток соприкасаются. Зоны контакта занимают 5% поверхности клеточной мембраны. Здесь происходит передача возбуждения от одной клетки к другой. Между гладкомышечными клетками вплетены волокна соединительной ткани, образующей плотный футляр (рис. 40) [191].
Советскими учёными А. А. Заварзиным, Н. Г. Хлопиным и др. установлено, что гладкие мышцы – группа различных по происхождению тканей, объединяемых единым функциональным признаком – способностью к сокращению. У позвоночных гладкие мышцы слюнных, потовых и молочных желёз происходят из эктодермы, а внутренних органов – из мезенхимы [79].
Гладкомышечные волокна обладают высокой чувствительностью к различным БАВ: адреналину, норадреналину, ацетилхолину, гистамину и др., а также к таким факторам, как РО2, РС02 и Ph. Это обусловлено наличием специфических рецепторов на их мембранах.
Сократительный аппарат гладких мышц не обладает столь упорядоченным строением, как в поперечнополосатых мышцах. Миофибриллы состоят из нитей актина и миозина, которые идут в различных направлениях и прикрепляющихся к разным участкам сарколеммы.
Укорачиваются гладкие мышцы также за счет взаимодействия актиновых и миозиновых нитей друг с другом. Однако расщепление АТФ, необходимое для этого процесса, происходит в 100 - 1000 раз медленнее, чем у поперечнополосатых мышц. Поэтому гладкие мышцы способны долго находиться в состоянии сокращения, затрачивая сравнительно мало энергии и не подвергаясь утомлению.
В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение изменяет мембранный потенциал клеток, увеличивает частоту потенциалов действия и в конечном итоге повышает её тонус. Кроме того, гладкомышечные клетки сами являются рецепторами растяжения.
В организме человека это свойство служит одним из способов регуляции двигательной деятельности внутренних органов. Например, увеличение тонуса желудка в ответ на растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. В кровеносных сосудах растяжение, создаваемое колебаниями кровяного давления, является основным фактором миогенной саморегуляции тонуса сосудов.
Еще одной специфической характеристикой гладкой мышцы является вязкость. Если её длительно удерживать в состоянии удлинения, то напряжение будет постепенно падать [240].
Таким образом, по своим основным свойствам – способности к сократимости (спонтанной или инициированной растяжением) и вязкости гладкие мышцы принципиально не отличаются от поперечнополосатых.
Цепной миотатический рефлекс
В 1905 г. немецкий биолог Якоб фон Икскюль (1864 – 1944) впервые продемонстрировал существование цепных миотатических рефлексов на офиурах (змеехвостках) – разновидности морских звезд. Их тело имеет звёздообразную форму с очень подвижными членистыми лучами, напоминающими хвосты змей. Лучи представляют собой ряд позвонков, соединённых мышцами (фото 15).

Фото 15. Морская звезда – офиура (змеехвостка)
Икскюль закреплял изолированный луч офиуры горизонтально в штативе, так, чтобы его конец свободно свисал, изогнувшись вниз, и наносил на него раздражение. При этом луч всегда отклонялся в сторону растянутых мышц, т.е. кверху [цит. по 134].
Подобный механизм координации описан также и у низших червей – немартин [16]. При раздражении переднего конца червя сначала сокращаются продольные мышцы головного отдела; затем постепенно, переходя от участка – к участку, волна сокращения достигает противоположного конца. Сокращение предыдущего участка ведет к растяжению ближайшего последующего сегмента тела. В растянутом сегменте раздражаются кожно-мышечные рецепторы, инициируя его сокращение.
Цепные координации поперечнополосатой мускулатуры обнаружены и у млекопитающих. Их существование было наглядно продемонстрировано Р. Магнусом [134] на хвосте децеребрированной, а затем спинализированной кошки.
Животное укладывали на стол спиной кверху, чтобы хвост свободно свисал через его край. При раздражении кончика хвоста он также всегда отклонялся вверх – в сторону растянутых мышц.
После того как переднюю часть туловища кошки перемещали в боковое положение, реакция хвоста менялась. Теперь при раздражении он двигался в сторону направления передних лап животного (рис. 27).

Рис. 27. Опыт Р. Магнуса по выявлению цепного миотатического рефлекса в хвосте децеребрированной и спинализированной кошки [134]
Перерезка спинного мозга нарушила связи передней части туловища с двигательными центрами хвоста, однако поворот тела вызывает асимметричное растяжение мышц с обеих сторон хвоста. Это способствовало изменению направления рефлекса.

Рис. 28. Образец миотатической цепи, включающей контрлатеральную наружную (1) и ипсилатеральную внутреннюю (2) косые мышцы живота; переднюю зубчатую (3), малую грудную (4) и межрёберные мышцы (5)
По данным И.С. Беритова [16], цепной миотатический рефлекс может развиться как при быстром, так и при медленном растяжении мышцы и сохраняется все время, пока действует растягивающая сила. Он может появляться при растяжении мышцы всего на 0,8 % ее длины покоя.
Достаточно взглянуть в учебник анатомии, чтобы убедиться в том, что субстраты для реализации цепных миотатических рефлексов существуют и у человека. Действительно, многие весьма крупные массивы скелетных мышц имеют одинаково направленную исчерченность рельефа. Нередко одна мышца как бы является продолжением другой. Эти цепочки мышц могут иметь диагональную или продольную ориентацию.
Например, волокна наружной косой мышцы живота, сохраняя преемственность по направлению, продолжаются через белую линию в волокна внутренней косой мышцы, а те, в свою очередь, плавно переходят в пучки брюшной части большой грудной и передней зубчатой мышцы (рис. 28). Последняя аналогичным образом связана с подлопаточной и ромбовидными мышцами.
В качестве другого примера можно привести цепочку, состоящую из икроножной мышцы, ишиокруральной мускулатуры и мышц ягодицы (рис. 29).

Рис. 29. Образец миотатической цепи, включающей икроножную мышцу (1), а также ишиокруральную (2) и ягодичную (3) мускулатуру
Сходство внутренней архитектуры весьма удаленных друг от друга мышц, имеющих различную иннервацию, может быть объяснено единством их функции на более ранних этапах эволюции. Возможно, некоторые из этих закономерностей отражают спиралевидно-волновой или удлинительно-укоротительный стереотипы движений, используемый червями, рыбами или змеями для перемещения в пространстве.
Клинико-экспериментальным обоснованием существования цепных мышечных координаций у человека могут послужить работы Т.Н. Несмеяновой и А.Н. Транквиллитати [152], которым в процессе обучения у больных с полным перерывом спинного мозга при наличии спастической параплегии удавалось добиться включения в двигательную активность мышц, иннервируемых сегментами, расположенными ниже уровня поражения.

Фото 16. Больная со спинальной параплегией, обученная сгибанию и разгибанию голени при помощи цепных миотатических синкинезий [152]
В этом случае запуск движений производился из расположенных выше уровня поражения верхних участков длинных мышц туловища и плечевого пояса, которые напрягались произвольно путем подтягивания на руках за изголовье кровати (фото 16). Далее с коротким интервалом последовательно сокращались парализованные мышцы: нижние отделы выпрямителя позвоночника, косые мышцы живота, средняя ягодичная мышца, четырехглавая и двуглавая мышцы бедра. В результате происходило слабое сгибание голени. Возникала как бы цепь миотатических рефлексов, в которой сокращение одной мышцы активирует дистально расположенную мускулатуру.



Рис. 30. Миофасциальные меридианы, выделяемые в Рольфинге [314].
Ориентация мышечных цепей: а – продольная, б – диагональная, в – латеро-латеральная
Не исключено, что у людей с интактной нервной системой эти древние связи также могут служить анатомическим субстратом для распространения мышечно-тонических и некоторых других реакций из очага первичной ирритации. Данный вид функциональных связей мы предлагаем называть термином «миотатические синкинезии». Вероятно при определённых условиях они могут иметь патогенетическое значение.
Можно предположить, что любое локальное напряжение мышцы, вызывая растяжение ее ближайших соседей по мышечной цепи, способствует активации миотатического рефлекса и повышению тонуса в последних. Таким образом, развивается цепной рефлекс, приводящий к последовательной тонизации всех мышц данной цепи, даже если они весьма удалены от очага первичного поражения. При этом сравнительно подвижные детали скелета (лопатка, ребра, тазовая кость, надколенник и т.д.) могут включаться в мышечную цепь как пассивные костные вставки, существенно не препятствуя распространению мышечно-тонических реакций [178].
Напряжение головной мышцы цепи, вероятно, инициируется позно-тоническими перегрузками или висцеро-моторными влияниями [53,230]. В дальнейшем миотатическая синкинезия может стабилизироваться за счёт формирования в её звеньях миофасциальных триггерных точек (ТТ). Согласно современным представлениям, последние имеют следующий патогенез.
Вследствие временных задержек в распространении двигательного нервного импульса мышечные волокна, расположенные в области брюшка мышцы, недалеко от мионеврального синапса, активируются раньше остальных. В результате, при любом сокращении мышцы имеется тенденция к растяжению ее более тонких и слабых участков, прилежащих к сухожилию. Особенно неблагоприятны в этом отношении мышцы со сложной внутренней архитектурой, например, перистые [193]. Поскольку мышце как биомеханическому телу свойственна вязкость, то после её расслабления изменение первоначальной конфигурации волокон может сохраниться, одни из них оказываются сжатыми, другие – растянутыми. Если в зону деформированного участка попадает мышечное веретено – создаются условия для искажения проприоцептивной импульсации и самоподдержания сегментарного миотатического рефлекса, что в конечном итоге и приводит к формированию ТТ [85].

Рис. 31. Схема формирования спиральных «миоадаптивных» цепей по Л.А. Кадыровой с соавт. [95]
По-видимому, первая попытка осознанного использования мышечных цепей с лечебной целью принадлежит врачу-остеопату Иде Рольф (1896–1979). В основу созданного ею направления – Рольфинга – было положено понятие о «миофасциальных меридианах», определяемых как типичные линии, по которым распространяется энергия напряжённых мышц (мышечная тяга). Эти «линии», соединяясь друг с другом, покрывают всё тело в виде сети. Различают цепи продольной, диагональной и латеро-латеральной ориентации (рис. 30). Каждый пациент имеет свой специфический набор клинически актуальных миофасциальных цепей, который может меняться от сеанса к сеансу [314].

Рис. 32. Образцы миофасциальных цепей по О.В. Кузнецову [116]
а – дорзальная поверхностная миофасциальная цепь, б – вен-тральная поверхностная миофасциальная цепь, в – спиральная миофасциальная цепь
Современные вертеброневрологи – Л. А. Кадырова и Н.Н. Сак с соавт. [96, 95, 209, 208] предполагают, что в скелетной мускулатуре человека могут быть выделены своеобразные мышечные цепи, оплетающие конечности и туловище наподобие спиралей (рис. 31). Распределение рефлекторной мышечной патологии находится в прямой зависимости от этих спиралевидных кинематических цепей.
Специалист по прикладной кинезиологии О.В. Кузнецов [116] утверждает, что на основании мануального мышечного тестирования [25, 302] можно выделить несколько вариантов миофасциальных цепей поверхностного и глубокого залегания, имеющих, как правило, диагональный или спиральный характер.
Например, в состав дорзальной цепи входят: подошвенная фасция стопы и короткие сгибатели пальцев, икроножная мышца, разгибатели бедра, крестцово-бугорная связка – на одной стороне, а также крестцово-поясничная фасция, выпрямитель позвоночника и апоневроз головы – на другой (рис. 32-а).
Вентральная мышечная цепь представлена коротким и длинным разгибателями пальцев, передней большеберцовой мышцей, надколенником и прямой головкой четырёхглавой мышцы бедра – на одной стороне и прямой мышцей живота, фасцией грудины, грудинно-ключично-сосцевидной мышцей и апоневрозом головы – на другой (рис. 32-б).
Спиральная цепь в области нижней конечностипредставлена передней большеберцовой, малоберцовыми мышцами и мышцей, напрягающей широкую фасцию бедра. Далее она диагонально продолжается по вентральной поверхности туловища в составе косых мышц живота той же стороны тела и передней зубчатой мышцы – противоположной (рис. 32-в).
Р. Смичек [218] полагает, что мышечные цепочки не является неизменными анатомическими образованиями. Они представляет собой временную группу анатомических единиц, которые формируются для стабилизации туловища во время движения или внешнего силового воздействия. Одни и те же мышцы могут входить в состав различных мышечных цепочек. Участие отдельных мышечных цепей различно даже при выполнении отдельных фаз движения.
По его мнению, слаженная работа мышечной цепи нарушается из-за слабости или функциональной неактивности отдельных её звеньев. Одной из причин такой дисфункции мышц может быть патологическая импульсация из поражённого позвоночно-двигательного сегмента (рис. 33).

Рис. 33. Варианты мышечных цепей по Р. Смичеку [218]