Учебная информация для использования на занятии
Эндокринная система – трёхуровневая, многокомпонентная, гуморально взаимосвязанная, оказывающая влияние на процессы развития, роста, размножения, обмен веществ, обеспечивающая постоянство внутренней среды и биоритмы.
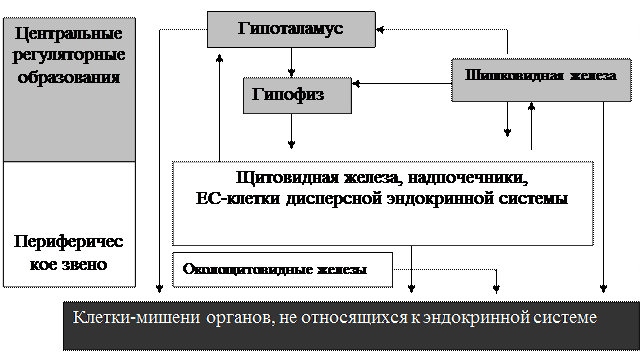
Рис. 1. Гуморальные взаимосвязи в эндокринной системе.
Гуморальная регуляция обусловлена сродством гормонов, вырабатываемых компонентами эндокринной системы, и рецепторов клеток-мишеней, изменяющих функциональное состояние под влиянием гуморальных сигналов. Эффект действия может быть стимулирующим (гипоталамуса на гипофиз, гипофиза и шишковидной железы на компоненты периферического звена) или угнетающим (гормонов шишковидной железы на гипоталамус и гипофиз, гипоталамуса на гипофиз, щитовидной и надпочечных желёз на гипоталамус). Низкая ночная активность ЕС-клеток стимулирует шишковидную железу. В цепи взаимосвязанных компонентов эффект усиливается в направлении от центра к периферии (акселерация), развивается постепенно и длительно сохраняется. Иерархия, акселерация и обратные взаимосвязи – основные принципы функционирования эндокринной системы (рис. 1).
Щитовидная железа – самая крупная, дольчатая, окружённая капсулой, общей для околощитовидных желёз, с эпителиальной паренхимой, организованной в фолликуллы и интерфолликулярно, развитой сетью кровеносных сосудов в соединительнотканной строме.
Развитие щитовидной железы осуществляется из общего эмбрионального зачатка для околощитовидных желёз и тимуса – передней (глоточной) кишки эмбриона. Органы составляют группу бранхиогенных желёз. Их зачатки у человека появляются на 3-4 неделях эмбриогенеза: щитовидной – непарный на уровне I-II пар жаберных (глоточных) карманов, околощитовидных – парные из III и IV пар карманов. Зачаток щитовидной железы приобретает вид протока (щитоязычного), который растёт вдоль вентральной поверхности кишки и на дистальном конце формирует две доли. К концу 12-й недели в долях дифференцируются фолликулярные клетки, формируются фолликулы, заполненные коллоидом, вступающие во взаимоотношение со стромой мезенхимной природы и нейробластами нервного гребня. Последние дифференцируются в парафолликулярные клетки. Функциональная активность щитовидной железы наиболее высокая в детском возрасте.
Фолликулярные клетки составляют 98% от общего количества клеток паренхимы, располагаются одним слоем на базальной мембране. Фолликулы – аденомеры железы обычно имеют округлую форму, диаметр 0,05-0,5 мм, заполнены оксифильным коллоидом. Который даёт положительную ШИК-реакцию на гликопротеиды.
В нормальном функциональном состоянии фолликулярные клетки кубической формы с округлым ядром, ядрышками, умеренно конденсированным хроматином и базофильной цитоплазмой. На апикальной поверхности имеются микроворсинки. В околоядерной области располагается гранулярная эндоплазматическая сеть (ЭПС), комплекс Гольджи, лизосомы, митохондрии, транспортные, секреторные и пиноцитозные везикулы.
В секреторном цикле фолликулярной клетки условно выделяют две фазы: продукции и выделения йодсодержащих гормонов.
В фазу продукции клетка синтезирует в гранулярной ЭПС белок, который в комплексе Гольджи гликозилируется и превращается в тиреоглобулин, транспортируемый в составе секреторных везикул к адлюминальной поверхности и путём экзоцитоза выделяющийся в полость фолликула. Стимулированная ТТГ, фолликулярная клетка извлекает из крови ионы йода, восстанавливает их с помощью фермента пероксидазы. Атомы йода йодируют тиреоглобулин в составе коллоида. Образуются монойодтиронин, дийодтиронин и их комплексы. Интрафолликулярно под воздействием ферментов, выделяемых лизосомами, осуществляется протеолиз йодированного тиреоглобулина с высвобождением трийодтиронина (Т3) и тетрайодтиронина (Т4), которые являются гормонами.
В фазу выведения гормоны поглощаются путём пиноцитоза, транспортируются в везикулах к базальной поверхности плазмолеммы и выделяются в кровь. Они необходимы для нормального роста, дифференцировки и функционирования головного мозга, поддержания основного обмена, температуры тела.
При дефиците йодсодержащих гормонов в детском возрасте развивается критинизм. У взрослых страдают умственная способность. Избыток гормонов способствует разобщению процессов окисления и фосфорилирования в митохондриях, стимулируя термогенез и развитие экзофтальма.
Недостаток йода в пище осложняет выработку йодсодержащих гормонов. По механизму отрицательной обратной связи в гипоталамусе активизируется секреция тиреолиберина, в гипофизе – ТТГ, под влияние которого развивается гиперфункция щитовидной железы. При гиперфункции фолликулярные клетки делятся, приобретают столбчатую форму с базальными складками и псевдоподиями на апикальной поверхности. Ядро становится овальным, цитоплазма – апикально зернистой, с частицами фагоцитированного коллоида. Количество и плотность (оксифилия) коллоида внутри фолликулов снижается, он как бы вспенивается. Фолликулы становятся складчатыми. Протеолиз осуществляется как в полости фолликулов, так и внутриклеточно при слиянии лизосом с фагосомами.
При стрессе в гипоталамусе угнетается продукция тиреолиберина. Активность тиротропоцитов гипофиза снижается. В щитовидной железе проявляется гипофункция: фолликулярные клетки уплощаются, их ядра становятся тёмными, адлюминальная и базальная поверхности гладкими, прекращается йодирование, расщепление тироглобулина и секреция в кровь Т3 и Т4 . Коллоид уплотняется, растягивает фолликулы.
Парафолликулярные клетки (кальцитониноциты, или С-клетки) составляют 0,1% от общего количества клеток паренхимы и не чувствительны к ТТГ. Располагаются в фолликулах между фолликулярными клетками. Имеют коническую форму, округлое ядро с ядрышками, осмиофильные секреторные гранулы, умеренно развитую гранулярную ЭПС, комплекс Гольджи, митохондрии. Преобладают более мелкие электронно-плотные секреторные везикулы, содержащие гормон пептидной природы кальцитонин. На плазмолемме имеются рецепторы, чувствительные к возрастанию содержания ионов Са2+ в крови. Клетки-мишени – остеобласты (кальцитонин стимулирует их Са2+–связывающую деятельность в костной ткани), остеокласты (подавляет резорбционную активность многоядерных клеток костной ткани), энтероциты кишечника (подавляет всасывательную активность) и эпителиоциты канальцев почек (повышает экскрецию Са2+). В результате содержания Са2+ в крови восстанавливается (гипокальциемическое действие).
Интерфолликулярные клетки составляют 2% клеток паренхимы щитовидной железы, образуют камбий.
Околощитовидная железа – самая мелкая (около 5 мм), парная (по две с каждой стороны), дольчатая с соединительнотканной стромой и эпителиальной паренхимой, которая состоит из анастомозирующих тяжей эндокринных клеток. С возрастом строма разрастается, в ней увеличивается количество жировых клеток. В паренхиме у 5-летних детей появляются крупные оксифильные паратироциты. Главные паратироциты имеют размеры 7-10 мкм, округлое ядро с ядрышками и подразделяются на светлые (20%) и тёмные (80%). Светлые – функционально неактивные, бедные органеллами с липидными каплями и гранулами гликогена. Тёмные рецепторами на плазмолемме улавливают снижение содержания Са2+ в крови, вырабатывают в гранулярной ЭПС гормон паратирин, в комплексе Гольджи оформляют его в секреторные везикулы диаметром 120-200 нм и по мерокринному способу выделяют в кровь, проявляя гиперкальциемический эффект. Паратирин стимулирует остеокласты, резорбирующие матрикс костной ткани. Оксифильные клетки имеют округлое ядро с ядрышком, многочисленные митохондрии, единичные цистерны гранулярной ЭПС, комплекс Гольджи и мелкие электронно-плотные секреторные гранулы, содержащие серотонин. Возможно, участвуют в регуляции суточной активности железы.
Надпочечник – парная железа размером 1х3х5 см с толстой капсулой из плотной соединительной ткани, под которой располагается корковое вещество, в центре – мозговое вещество.
Развитие надпочечников начинается на 5-й неделе эмбриогенеза человека с закладки из материала вентральной мезодермы первичной коры, состоящей из крупных оксифильных клеток, вторичной коры, образованной мелкими базофильными клетками, и мезенхимной капсулы и стромы. Из ганглиозной пластинки на 6-й неделе мигрируют клетки симпатогонии, которые на 7-й неделе обнаруживаются в капсуле, к 6-му месяцу проникают через корковое вещество в центральную область, формируя мозговое вещество. У новорождённых первичная кора претерпевает инволюцию (в течение 2-х недель – на 50%). Во вторичной коре, расположенной под капсулой железы, у детей начинает проявляться зональность. Вторичная кора и мозговое вещество разрастаются, достигая в пубертатном возрасте параметров, характерных для взрослых людей.
Корковое вещество составляет 70-80% объёма органа и подразделяется на зоны – клубочковую, пучковую и сетчатую в соотношении (у взрослых людей) 3:15:2. Паренхима состоит из тяжей эндокринных клеток кортикостероцитов. В узких прослойках стромы располагаются капилляры с фенестрированным эндотелием, которые на границе с мозговым веществом отдают кровь сосудам синусоидного типа. В клубочковой зоне тяжи клеток, подворачиваясь под капсулу, формируют округлые аркады, напоминающие клубочки. В пучковой зоне они следуют радиально, в сетчатой несколько разрыхляются и анастомозируют.
Кортикостероциты клубочковой зоны, чувствительные к АГТГ шишковидной железы, конической формы высотой до15 мкм с округлым ядром, ядрышками и слабо воспринимающей красители цитоплазмой. Содержат агранулярную ЭПС, митохондрии, жировые включения. Из холестерина, запасаемого в липидных каплях, в агранулярном ретикулуме синтезируются минералокортикоиды – гормоны стероидной природы (альдостерон, кортиксон), регулирующие электролитный обмен. Основной гормон – альдостерон, предотвращающий в почках потерю ионов Na+ с мочой и способствующий увеличению артериального давления.
Кортикостероциты узкой наружной части пучковой зоны – недифференцированные кубической формы размером 8-10 мкм с округлым ядром, ядрышками, бедной органеллами и не содержащей липидных капель цитоплазмой. Составляют суданофобную прослойку. Стимулированные АКТГ гипофиза, делятся, оттесняются в пучковую и сетчатую зоны, участвуют в обновлении паренхимы, клетки которой в данных зона также чувствительны к АКТГ.
Кортикостероциты широкой внутренней части пучковой зоны – самые крупные, до 25 мкм, губчатые (теряют липиды при проводке через спирты, при окраске замороженных срезов суданом III жировые включения окрашиваются в оранжевый цвет). Имеют митохондрии с тубулярными кристами. В гладкой ЭПС синтезируют глюкокортикоиды кортизол, гидрокортизол, кортикостерон, регулирующие углеводный, белковый, жировой обмен, угнетающие иммунную систему и развитие воспалительного процесса. Гормоны в цитоплазме не накапливаются. При гиперфункции число жировых капель уменьшается.
Кортикостероциты сетчатой зоны кубической формы размером 15-20 мкм с округлым ядром и оксифильной цитоплазмой. Содержат митохондрии с тубулярными кристами, гладкую ЭПС, незначительное количество жировых включений, синтезируют половые гормоны (основной – дегидроэпиандростерон, стимулирующий развитие вторичных половых признаков мужского пола). Сетчатая зона является «кладбищем» коры.
Мозговое вещество имеет паренхиму гроздьевидного типа. Скопления хромаффинных его клеток разделяют широкие прослойки соединительной ткани, содержащие крупные синусоидные капилляры и вены. Мультиполярные вегетативные нейроны, стимулирующие деятельность клеток паренхимы мозгового вещества, окружают мантийные глиоциты. Хромаффинные клетки вырабатывают катехоламины, восстанавливающие бихромат калия (бурая окраска), ростовые факторы и гормоны пептидной природы. Различают светлые хромаффинные клетки, вырабатывающие адреналин (адреналоциты), и тёмные, вырабатывающие норадреналин (норадреналоциты), проявляющий более выраженные восстанавливающие свойства. Ядра клеток округлые светлые с ядрышками, митохондрии мелкие, гранулярная ЭПС и комплекс Гольджи развиты, секреторные гранулы диаметром 150-500 нм с электронно-плотной сердцевиной и светлым ободком.