История открытия биопотенциалов
| Первые попытки по изучению биоэлектрических явлений («животного электричества») известны с ХVIII века, когда были выполнены исследования на «электрических» органах рыб (Адансон, 1751; Целп, 1773; Вильямсон, 1775 и др.). Все эти исследования подготовили благоприятную почву для трудов Гальвани, заложивших основу электрофизиологии как вполне самостоятельной области науки. В 1791 г. им были опубликованы результаты исследований, в том числе знаменитого «балконного» опыта (Рис. 1.5). |
|
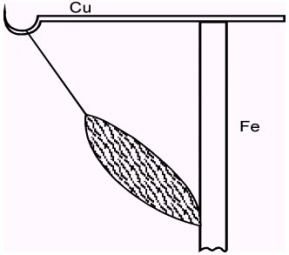
| Рис.1.5 Балконный опыт Гальвани (по Латманизовой Л.В.). |
| При подвешивании нервно - мышечного препарата на железную решетку с помощью медного крючка, проходящего через спинной мозг препарата, имело место сокращение мышц лапки каждый раз, когда эта лапка соприкасалась с железной решеткой балкона. |
| Гальвани считал, что причиной сокращения мышцы в данном случае является электричество, причем, источник этого электричества ученый видел именно в нервно - мышечном препарате лягушки. |
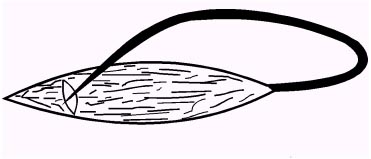
| Однако ему возразил его знаменитый соотечественник - физик А. Вольта, который считал, что в «балконном» опыте мышца является лишь чувствительным «электрометром» электричества, порождаемого контактной разностью потенциалов разных металлов, используемых в опытах Гальвани. Позднее, защищая свою точку зрения от возражений оппонентов, Гальвани воспроизводит различные модификации опытов, в которых сокращение мышцы вызывалось путем набрасывания нерва с помощью стеклянной палочки на поврежденный и неповрежденный участок мышцы (рис.1.6). |
|
| Рис.1.6 Модификация «балконного» опыта Гальвани. |
| Позднее открытия Гальвани были подтверждены в работах Маттеучи (1837). Однако Маттеучи обнаружил явление вторичного или индуцированного сокращения: при помещении нерва одного нервно-мышечного препарата на мышцу другого препарата и раздражителя нерва этого препарата, Маттеучи наблюдал сокращение мышцы обоих препаратов (рис.1.7). |

|
| Рис.1.7. Опыт Маттеучи: вторичное (индуцированное) сокращение мышцы. |
| На основании этого явления Маттеучи выдвинул предположение об изменении электрических зарядов нервной ткани при ее возбуждении. |
| Дальнейшее развитие представлений о природе «животного электричества» связано с внедрением в физиологию экспериментальных приемов и техники. В 1820 году Швейгер сконструировал гальванометр, усовершенствовав который итальянский физик Нобиле применил его в 1827 г. для проверки опытов Гальвани. Однако наибольший интерес представляют работы Э.Дюбуа-Реймона, выполненные в 1840-1860 гг. В этих работах благодаря высокочувствительному гальванометру и ряду других технических новшеств удалось впервые определив электрические процессы в мышце, зарегистрировав потенциал наружной и внутренней поверхности мембраны клеток. Впервые он установил, что наружная мембрана заряжена положительно по отношению к внутренней, и эта разность потенциалов изменяется при сокращении мышцы. |
| Позднее, в 1896 г. В.Ю. Чаговец впервые высказал гипотезу о ионном механизме электрических потенциалов в живых клетках и сделал попытку применить для их объяснения теорию электролитической диссоциации Аррениуса. В 1902 г. Бернштейном была развита мембранно-ионная теория, согласно которой клеточная поверхность представляет собой полупроницаемую мембрану, которая в состоянии физиологического покоя проницаема для ионов калия и практически непроницаема для остальных вне - и внутриклеточных ионов. |
| В 1936 году английский зоолог Джон Юнг обнаружил у кальмаров и каракатиц необычайно толстые аксоны, которые впоследствии стали называть "гигантскими аксонами". Их диаметр превышал 0,5 мм, что позволило достаточно легко вводить в них микроэлектроды, проводить химический анализ содержащейся в них жидкости, вводить в них различные растворы и т.д. «Гигантские аксоны» стали излюбленным объектом для изучения биоэлектрических явлений в тканях, с их помощью было получено много новых и интересных данных. |
| Современные представления о природе биоэлектрических явлений в тканях базируются на результатах работ Алана Ходжкина, Эндрью Хаксли, Бернарда Катца. Эти ученые в 40-50 годах нашего века модифицировали и экспериментально обосновали мембранно-ионную теорию Ю. Бернштейна. В настоящее время их взгляды о природе биоэлектрических явлений пользуются всеобщим признанием. Согласно их представлениям, наличие электрических потенциалов в живых клетках обусловлено различной концентрацией ионов Na+, K+, Ca2+ и Cl- внутри и вне клетки, а также различной проницаемостью для них клеточной мембраны. За разработку теории ионного механизма возбуждения эти авторы были удостоены звания лауреатов Нобелевской премии. |
Общепринятой теорией возникновения биопотенциалов является мембранно-ионная теория. Согласно мембранно- ионной теории, причиной возникновения разности потенциалов - это неравномерное распределение ионов по обе стороны клеточной мембраны (в системе цитоплазма - окружающая среда). Авторы этой теории: В.Ю. Чаговец - 1896 г., Бернштейн - 1902-1903 гг., Ходжкин, Хаксли, Кац -1940 -1941 гг.
Краткая суть теории состоит в следующем :
- биотоки, биопотенциалы возникают толь ко в возбудимых тканях (нервная, мышечная, железистая).
- клетки возбудимых тканей имеют особо устроенные биологические мембраны, в которых находятся ионные «ворота», ионные «насосы».
- что внутри клетки и за ее пределами (внеклеточное пространство) всегда имеется концетрационный градиент ионов калия и натрия (калия больше внутри клетки, а натрия больше во внеклеточной среде).
- что ионные каналы динамичны, и их открытие зависит от функционального состояния клетки или ткани (покой или возбуждение)
-что в состоянии покоя открыты калиевые каналы, а в состоянии возбуждения - натриевые каналы.
- что в состоянии покоя клетка генерирует потенциалы покоя, а в состоянии возбуждения –потенциалы действия .
Таким образом, биоэлектрические явления в тканях - это разность потенциалов, которая возникает в тканях в процессе нормальной жизнедеятельности. Эти явления можно регистрировать, используя трансмембранный способ регистрации. При этом один электрод располагается на наружной поверхности клетки, другой - на внутренней.
При таком способе регистрируются:
1.потенциал покоя или мембранный потенциал;
2.потенциал действия.