Физиологическая характеристика возбудимых тканей
Возбудимость как высшее свойство возбудимых тканей проявляется при действии факторов внешней или внутренней среды, называемых раздражителями. Различают две группы раздражителей:
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т. п.), физико-химические (осмотические – кристаллик хлорида натрия).
Классификация раздражителей по биологическому принципу:
1) адекватные, которые при минимальных энергетических затратах вызывают возбуждение ткани в естественных условиях существования организма;
2) неадекватные, которые вызывают в тканях возбуждение при достаточной силе и продолжительном воздействии.
К общим физиологическим свойствам возбудимых тканей относятся:
1) возбудимость– способность живой ткани отвечать на действие достаточно сильного, быстрого и длительно действующего раздражителя изменением физиологических свойств и возникновением процесса возбуждения.
Мерой возбудимости является порог раздражения. Порог раздражения– это та минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции. Так как порог раздражения характеризует и возбудимость, он может быть назван и порогом возбудимости. Раздражение меньшей интенсивности, не вызывающее ответные реакции, называют подпороговым;
2) проводимость– способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани;
3) рефрактерность– временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель);
4) лабильность– способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации.
Существует определенная зависимость ответной реакции от параметра раздражителя.
Эта зависимость характерна для возбудимых тканей и выражается в трех следующих закономерностях:
1) закон силы раздражения;
2) закон длительности раздражения;
3) закон градиента раздражения.
Закон силы раздраженияустанавливает зависимость ответной реакции от силы раздражителя. Ответная реакция ткани пропорциональна силе наносимых раздражений до определенного предела. Увеличение ответной реакции - результат возбуждения все большего числа волокон ткани. При действии максимального раздражителя возникает наибольшая ответная реакция, т. к. все волокна вовлечены в процесс возбуждения и дальнейшее увеличение ответной реакции невозможно.
Эта зависимость неодинакова для отдельных клеток и для целой ткани. Для одиночных клеток зависимость называется «все или ничего». Характер ответной реакции в данном случае зависит от достаточной пороговой величины раздражителя. При воздействии подпороговой величиной раздражения ответной реакции возникать не будет (ничего). При достижении раздражения пороговой величины возникает ответная реакция, она будет одинакова при действии пороговой и любой сверхпороговой величины раздражителя (часть закона – все).Это характерно, например, для сердечной мышцы, являющейся сплошным синцитием, а также для одиночного мышечного волокна.
Для совокупности клеток (для ткани) эта зависимость иная, ответная реакция ткани прямо пропорциональна до определенного предела силе наносимого раздражения. Увеличение ответной реакции связано с тем, что увеличивается количество структур, вовлекающихся в ответную реакцию.
Закон длительности раздражений.
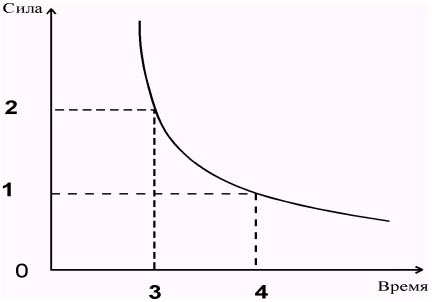
| Величина ответной реакции возбудимых тканей прямо пропорциональна (в определенных пределах) силе раздражителя эта зависимость выражается уравнением: В = 1/S, где В - возбудимость, S - сила раздражителя. Однако не только сила раздражителя, но и время (длительность) его воздействия оказывает влияние на величину ответной реакции. Более того, пороговый раздражитель при более коротком времени воздействия может не вызвать ответной реакции, т.е. оказаться допороговым. Представленная на (рис. 1.4) кривая называется кривой силы - длительности, или силы - времени. Она была получена при исследовании возбудимости нервов и мышц Горвегом (1892), Вейсом (1901) и Лапиком (1909). |
|
| Рис. 1.4. Зависимость между силой и длительностью порогового раздражения: 0-1 - реобаза, 0-3 - хронаксия, 0-4 - полезное время, 0-2 две реобазы; по оси абсцисс - длительность раздражения, по оси ординат - сила раздражения (в усл. ед.) |
| По этой кривой можно судить о том, что раздражитель ниже некоторой минимальной силы не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой (ордината 0-1). Наименьшее время (отрезок 0-4), в течение которого должен действовать раздражитель силой в одну реобазу, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. Как видно, при очень коротких стимулах кривая силы - времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражителя. Поэтому, кроме полезного времени, в качестве меры времени раздражения Лапик ввел понятие "хронаксия". Хронаксия - это время, в течение которого должен действовать ток равный двум реобазам, чтобы вызвать возбуждение. |
| Как видно, при очень коротких стимулах кривая силы - времени становится парралельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждение не возникает, как бы не велика была сила раздражителя. Поэтому, кроме полезного времени, в качестве меры раздражения Лапик ввел понятие «хронаксия». Хронаксия - это время, в течении которого должен действовать ток, равный двумя реобазам, чтобы вызвать возбуждение. |
Ответная реакция ткани зависит от длительности раздражения, но осуществляется в определенных пределах и носит прямо пропорциональный характер. Существует зависимость между силой раздражения и временем его действия. Эта зависимость выражается в виде кривой силы и времени и называется кривой Горвега—Вейса—Лапика. Кривая показывает, что каким бы сильным ни был бы раздражитель, он должен действовать определенный период времени. Если временной отрезок маленький, то ответная реакция не возникает. Если раздражитель слабый, то бы как длительно он ни действовал, ответная реакция не возникает. Сила раздражителя постепенно увеличивается, и в определенный момент возникает ответная реакция ткани. Эта сила достигает пороговой величины и называется реобазой (минимальной силой раздражения, которая вызывает первичную ответную реакцию). Время, в течение которого действует ток, равный реобазе, называется полезным временем.
Закон градиента раздражения. Градиент– это крутизна нарастания раздражения. Ответная реакция ткани зависит до определенного предела от градиента раздражения. При сильном раздражителе примерно на третий раз нанесения раздражения ответная реакция возникает быстрее, так как она имеет более сильный градиент. Если постепенно увеличивать порог раздражения, то в ткани возникает явление аккомодации. Аккомодация – это приспособление ткани к медленно нарастающему по силе раздражителю. Это явление связано с быстрым развитием инактивацииNa-каналов. Постепенно происходит увеличение порога раздражения, и раздражитель всегда остается подпороговым, т. е. порог раздражения увеличивается.
Законы раздражения возбудимых тканей объясняют зависимость ответной реакции от параметров раздражителя и обеспечивают адаптацию организмов к факторам внешней и внутренней среды.
Вывод:
- в зависимости от силы, длительности и градиента раздражителя наблюдается разная ответная реакция ткани;
2.эта зависимость не беспредельна.
Действие постоянного тока на ткани:
В 1859 г. Пфлюгер сформулировал закон полярного действия тока: при действии постоянного тока возбуждение возникает только в момент замыкания или только в момент размыкания цепи постоянного тока. При этом в момент замыкания оно возникает под катодом , а в момет размыкания - под анодом. ЭТО ЗАКОН ЯВЛЯЕТСЯ ПРОБНЫМ КАМНЕМ ОБЩЕЙ ФИЗИОЛОГИИ ВОЗБУДИМЫХ ТКАНЕЙ, А САМО ЭТО ЯВЛЕНИЕ - ФИЗИОЛОГИЧЕСКИМ ЭЛЕКТРОТОНОМ. Призамыкании цепи постоянного тока изменяется сила раздражителя. Поэтому возбуждение возникает либо в момент замыкания, либо в момент размыкания( по сути это закон градиента). При замыкании под катодом происходит деполяризация , и если она достаточна для достижения КУД , то возникает возбуждение – генерируется ПД. Если этого не происходит, то в области катода изменяется возбудимость: она возрастает вначале (анэлектротон, катэлектротон), но потом , вследствие инактивации натриевых каналов , возбудимость резко снижается. Это получило название катодической депрессии. (Вериго).Под анодом при замыкании цепи постоянного тока повышается величина мембранного потенциала – возникает гиперполяризация. Поэтому при замыкании цепи тока возбуждение не возникает. Возбудимость снижается(явление анэлектротона). Однако, если ток действует долго, то в условиях гиперполяризации КУД снижается (МП пирлижается к исходному уровню мембранного потнециала), поэтому при размыкании цепи тока в области анода возникает возбуждение. В целом, закон полярного действия тока может использоваться и в практике: если требуется заблокировать проведение возбуждения по нерву(болевая рецепция), то можно использовать постоянный ток, при этом в области расположения анода возбудимость будет снижена, что приведет к блоку проведения возбуждения. Аналогично – при длительной деполяризации в области катода ( когда возникает кактодическая депрессия).
Физиология нервных волокон, закономерности проведения возбуждения по нервным волокнам:
Нервные волокна _ это анатомические образования, состоящие из аксонов нервных клеток, покрытых общей оболочкой.Они обладают следующими физиологическими свойствами – Возбудимость, проводимость, рефрактерность, лабильность. В тоже время нервные волокна не являются самостоятельными структурными элементами нервной ткани, представляют собой комплеснообразование,включающее следующие элементы:отростки нервных клеток(аксоны)- осевые цилиндры, глиальные клетки, соединительнотканную (базальную) пластинку.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По ососбенностям строения и функциям нервные волокга подразделяются на два вида: миелиновые и безмиелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр составляет 5 – 7 мкм , а скорость проведения импульса 1 – 2 м/с. Миелиновык волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и аксоплазму. Миелиновая оболочка сосотит из липидов(80%), обладающих высоким омическим сопротивленим и на 20% -из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются перехватами Ранвье. Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12 – 20 мкм скорость проведения возбуждения составляет 70 – 120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа6 А, В и С.
Наибольшей скоростью проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с.Группа В имеет скорость от 3 до 14 м/с, а группа С – от 0,5 до 2м/с.
еПроцессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.