Типы химических связей. Природа ионизации, константа и степень ионизации. Различия в ионизации, обеспечивающие избирательность действия ксенобиотиков.
Оказавшись в непосредственной близости от места связывания (рецептора), молекула вещества немедленно вступает во взаимодействие. Длительность и избирательность этого взаимодействия определяется особенностями химической природы вещества и рецептора, а также типом связей между ними.
Рассмотрим наиболее важные типы химической связи: ковалентные, ионные (электростатические), водородные и вандерваальсовы. Эти основные типы могут подразделяться на множества отдельных разновидностей. Образование или разрыв любой из этих связей представляет собой истинную химическую реакцию, протекающую с заметным изменением энергии.
Прочность связей, обычно определяемую экспериментально, можно рассчитать, используя квантово-химические методы.
Ковалентная связь образуется за счет обобществления двумя атомами пары электронов, принадлежащих этим атомам. Она обычно значительно прочнее остальных. Прочность химической связи можно оценить по ее энергии.
Действие многих веществ можно прекратить простым отмыванием. Подобная быстрая обратимость означает, что проявление биологического эффекта не связано с образованием ковалентной связи.
Электростатические взаимодействия. Связи, образованные электростатическими силами, играют важную роль при взаимодействии веществ с мембранными структурами, точно так же, как при реакции субстратов с ферментами. Другая характерная особенность — легкость обмена ионов. Наиболее часто электростатические связи возникают между ионами (отсюда их название «ионные»).
Водородные связи образуются лишь при очень малом расстоянии между взаимодействующими атомами и достаточно строго ориентированы в пространстве, поэтому они обладают высокой избирательностью и направленностью, что очень важно при связывании вещества с рецептором. Кроме того, они играют основную роль в стабилизации конформаций молекул белков и нуклеиновых кислот.
Ван-дер-ваальсовы связи могут возникать только в тех случаях, когда геометрия двух молекул дает возможность двум атомам, способным к образованию связи, подойти друг к другу на достаточно близкое расстояние. Наличие в молекуле ксенобиотика ионизированной группы способствует сближению молекул до расстояния, на котором начинают действовать ван-дер-ваальсовы силы. Примером таких связей может служить взаимодействие между антигеном и его антителом, которое происходит исключительно за счет сил с малым радиусом действия, т. е. ван-дер-ваальсовых и водородных связей. Ван-дер-ваальсовы связи образуются благодаря тому, что все молекулы обладают энергией, достаточной для колебаний их атомов. Гидрофобные связи имеют большое значение при взаимодействии неполярных молекул ксенобиотиков с клеточными и внутриклеточными мембранами, для образования их комплексов с белками, при этом возможно нарушение конформации макромолекул.
В процессе взаимодействия низкомолекулярного вещества с биомишенью практически всегда формируется несколько типов связей, поскольку молекула токсиканта, как правило, включает полярный, и неполярный фрагменты.
Ион-дипольная связь. Многие неионизированные вещества имеют очень большие дипольные моменты. Такие молекулы могут притягиваться ионами и образовывать с ними связь. Ион-дипольные связи слабее ионной связи.
Большинство биологически активных веществ представляют собой кислоты, основания и соли и, следовательно, являются электролитами. Все электролиты понижают температуру замерзания воды в значительно большей степени, чем можно было бы ожидать исходя из их молярной концентрации.
Реально степень ионизации в растворе определяется только двумя факторами: рН раствора и рКа кислоты или основания.рКа (константа кислотности, или константа ионизации) является постоянной величиной для каждой кислоты или основания. Поэтому при определенной величине рН степень ионизации зависит только от природы кислоты, при этом не важно, были они нейтрализованы или нет.
Для описания состояния ионного равновесия Аррениус применил закон действующих масс. Так, уксусная кислота (СН3СООН) - это слабая кислота, ионизирующаяся в воде с образованием некоторого количества ионов водорода (Н+) и ацетат-ионов (СН3СОО):
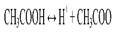
Соотношение произведения этих ионов [Н ][СН3СОО ] к концентрации неионизированных молекул [СН3СООН] всегда является постоянной величиной и определяет константу кислотности Ка или просто константу ионизации данной кислоты:
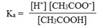
|
Степень ионизации любого вещества можно рассчитать при известных величинах рН раствора и рКа с помощью выражений:


Из данных уравнений следует, что зависимость степени ионизации от рН не является линейной, а выражается сигмоидной кривой.
В зависимости от степени ионизации ксенобиотики обладают различной биологической активностью и их можно разделить на три большие группы:
1ксенобиотики, обладающие большей биологической активностью в ионизированном состоянии;
2ксенобиотики, более активные в неионизированном состоянии;
3ксенобиотики, проявляющие биологическое действие в виде ионов и неионизированных молекул.
7. Вещества, обладающие большей биологической активностью в ионизированном состоянии. Вещества, менее активные в ионизированном состоянии и вещества, проявляющие биологическое действие в виде ионов и неионизированных молекул. Вещества, обладающие большей биологической активностью в ионизированном состоянии.
В началеXX в. было обнаружено, что многие органические катионы обладают антибактериальной активностью. Так алифатические амины, существующие при рН 7 главным образом в виде катионов, проявляют бактерицидное действие в отличие от ароматических аминов, которые существуют при рН 7 в основном в виде неионизированных молекул и таким действием не обладают.
Тогда антибактериальная активность подобных антисептиков объяснялась щелочностью их водных растворов. Однако при этом не учитывалось то обстоятельство, что те же основания, находящиеся в забуференном растворе, также обладают бактерицидными свойствами. Позднее выяснилось, что один из аминоакридинов (антибактериальные средства) — профлавин, или 3,6-диаминоакридин токсичен по отношению к широкому ряду грамположительных и грамотрицательных бактерий, но безвреден для тканей человека. Исследования серии аминоакридинов, проведенные Альбертом с сотрудниками в 1941 г., показали, что существует количественная связь между их антибактериальным действием и степенью ионизации по катионному типу.
Ксенобиотики, более активные в неионизироеаннот состоянии. Неионизированные вещества могут обладать очень сильным физиологическим действием (эфир, хлороформ и др.). Пример: ингибирования деления клеток яиц иглокожих салициловой кислотой.
В 1921 г. Вермст обнаружил, что многие слабые кислоты наиболее полно проявляют физиологическую активность в наименее ионизированном состоянии. Исследования проводились в интервале рН от 5 до 8. Степень (количество) ионизированных молекул растет в направлении увеличения рН. Оказалось, что кислота наиболее активна при рН 5. В этом случае количество неионизированной салициловой кислоты (рКа 3,0) составляет 0,99 %, что значительно больше, чем при остальных исследуемых значениях рН. Самое простое объяснение заключается в том, что нейтральные молекулы, а не анионы, салициловой кислоты ингибируют деление клеток.
При изучении действия слабых кислот на биологические объекты доказано, что количество вещества, необходимое для достижения эффекта, остается постоянным независимо от рН среды при условии, что рН по меньшей мере на одну единицу ниже, чем рКа; в этом случае не происходит ионизации токсического агента
В других опытах было также доказано, что эффективность слабых оснований возрастает с повышением рН, т. е. пропорционально снижению их ионизации, эффективность слабых кислот повышается при уменьшении рН, что также связано с уменьшением ионизации.
Ксенобиотики, проявляющие биологическое действие в виде ионов и неионизированныхмолекул. Многие вещества, особенно со значениями рКа в пределах от 6 до 8, проникнув в клетки в виде неионизированных молекул, проявляют свое биологическое действие как ионы. Например, доказано, что поступление бензойной кислоты в клетки дрожжей обратно пропорционально степени ионизации.
Ряд азотсодержащих гетероциклов, ионизирующихся с образованием аниона, сильно ингибируют реакцию Хилла при фотосинтезе. Однако было обнаружено, что при слишком высокой степени ионизации активность этих соединений теряется полностью. Это явление объясняется проникновением ингибиторов через мембрану в виде неионизированных молекул, а затем действием их как анионов. Такой механизм характерен для некоторых гербицидов.
Степень ионизации (образование анионов) многих противовоспалительных средств положительно коррелирует с их биологическим действием, если ионизации подвергаются не все молекулы и их липофильности достаточно для достижения места действия. Известно много ксенобиотиков, биологическая активность которых определяется действием неионизированных молекул и ионизированной части вещества.
При изучении биологической активности слабой кислоты обычно оказывается, что для получения стандартной реакции необходимо одинаковое количество ксенобиотика при любых значениях рН, которые на единицу и более ниже значения рКа. В этих условиях кислота слабо ионизирована и ее биологическая активность определяется прежде всего ионизированными молекулами. Таким образом, среди последней группы ксенобиотиков следует различать вещества, проявляющие биологическую активность в виде ионов и неионизированных молекул, и вещества, проходящие через мембрану в неионизированном состоянии и вызывающие биологический эффект внутри клеточного компартмента в виде ионов. На взаимодействие ксенобиотиков с мембраной оказывает влияние и ионизация мест связывания. Катионные препараты связываются с анионными центрами, которые могут иметь значения рКа от 2 до 7 (при наличии фосфатных групп), от 2 до 6 (при наличии карбоксильных групп) или 10 (в присутствии остатков тирозина, пиримидина, цистеина). Катионные центры должны иметь значения рКа 4 (аденин), 7 (гистидин), 10 (лизин), 13 (аргинин).
8. Проницаемость мембран для различных веществ. Транспорт ксенобиотиков через биологические мембраны: пассивная и облегченная диффузия, активный транспорт, пиноцитоз и фагоцитоз.
Физиологическая активность ксенобиотиков в значительной степени зависит от способности взаимодействовать с клеточными мембранами; не менее важное значение имеет их способность проникать внутрь клетки.
В настоящее время выделяют следующие формы прохождения ксенобиотиков (химических соединений) через мембраны:
1пассивная диффузия;
2облегченная диффузия (переносчики);
3активный транспорт веществ, выполняемый молекулярными машинами (АТФазы) и редокс-цепями (РЦ);
4активный транспорт веществ, осуществляемый за счет сопряжения термодинамических градиентов.
При рассмотрении основных механизмов переноса веществ через биологические мембраны различают пассивный и активный транспорт. При пассивной диффузии ксенобиотики проходят через мембрану в результате случайного молекулярного движения, и величина потока линейно зависит от концентрации и коэффициента проницаемости мембраны для данного вещества. Облегченная диффузия. Вещества, нерастворимые в липидах, с размером молекулы более 0,3-0,4 нм, не диффундируют через мембрану. Однако глюкоза и некоторые другие моносахариды с такими же размерами молекул, как правило, хорошо проникают в клетку. Для объяснения высокой проницаемости клеточных мембран по отношению к сахарам, аминокислотам и некоторым другим соединениям была выдвинута концепция «переносчиков», связывающихся с транспортируемым веществом и таким образом «облегчающих» их прохождение через мембрану. Такая облегченная диффузия происходит по градиенту концентрации без затрат энергии и относится к пассивному транспорту.
В основе механизма облегченной диффузии лежит обратимое соединение транспортируемого вещества со специфическим переносчиком, и образующийся комплекс вещество-переносчик диффундирует внутри мембраны от наружной поверхности к внутренней, где комплекс диссоциирует с высвобождением вещества внутрь клетки. Свободный же переносчик диффундирует назад к наружной поверхности мембраны, где соединяется с новой молекулой вещества, и цикл повторяется.
Активный транспорт веществ либо осуществляется за счет сопряжения электрохимических градиентов, либо выполняется молекулярными машинами (АТФазами). Активный перенос происходит с затратой энергии и идет против градиента электрохимического потенциала. Следует различать первичный и вторичный (или сопряженный) активный транспорт.
Первичный активный транспорт - трансмембранный векторный перенос веществ осуществляется непосредственно в ходе реакции энергетического преобразования в АТФазных системах или ОВЦ, т. е. используется энергия либо АТФ, либо энергия ОВ реакций. Вторичный активный перенос совершается, когда в качестве энергетических источников используются градиенты электрохимических потенциалов других ионов.
Следует отметить попадание в клетку веществ с помощью пиноцитоза и фагоцитоза. Транспортные белки (переносчики, АТФазы) способствуют проникновению через клеточные мембраны многих полярных молекул небольшого размера, однако они не способны транспортировать макромолекулы, например белки, полинуклеотиды, полисахариды, а также твердые частицы. Тем не менее в большинстве клеток указанные вещества проходят в обоих направлениях через плазматические мембраны. Механизмы, с помощью которых осуществляются эти процессы, сильно отличаются от механизмов, опосредующих транспорт небольших молекул и ионов. Пиноцитоз подразделяется на несколько этапов:
1) адсорбция на мембране молекул вещества; 2) впячивание или выпячивание (инвагинация) мембраны, образование пиноцитозного пузырька и отрыв его от мембраны с затратой энергии АТФ; 3) миграция пузырька внутрь протопласта, органеллы или наружу; 4) растворение мембраны пузырька (при действии фермента) или просто ее разрыв.