Хищничество. Механизмы стабилизации численности в системе хищник-жертва.
Хищничество — форма трофических взаимоотношений между организмами разных видов, при которых один из них (хищник) атакует другого (жертву) и питается его плотью, то есть обычно присутствует акт умерщвления жертвы.
Система «хищник — жертва» — сложная экосистема, для которой реализованы долговременные отношения между видами хищника и жертвы, типичный пример коэволюции.
Коэволюция — совместная эволюция биологических видов, взаимодействующих в экосистеме.
Отношения между хищниками и их жертвами развиваются циклически, являясь иллюстрацией нейтрального равновесия.
1. Единственным лимитирующим фактором, ограничивающим размножение жертв, является давление на них со стороны хищников. Ограниченность ресурсов среды для жертвы не учитывается.
2. Размножение хищников ограничивается количеством добытой им пищи (количеством жертв).
По своей сути модель Лотки–Вольтерры есть математическое описание дарвинского принципа борьбы за существование.
Система Вольтерра-Лотка, которую часто называют системой хищники-жертвы, описывает взаимодействие двух популяций - хищников (например, лисиц) и жертв (например, зайцев), которые живут по несколько разным "законам". Жертвы поддерживают свою популяцию за счет поедания природного ресурса, например, травы, что приводит к экспоненциальному росту численности популяции, если нет хищников. Хищники поддерживают свою популяцию за счет только "поедания" жертв. Поэтому, если популяция жертв исчезает, то вслед за этим популяция хищников экспоненциально убывает. Поедание хищниками жертв наносит ущерб популяции жертв, но в то же время дает дополнительный ресурс к размножению хищников.
Вопрос
ПРИНЦИП МИНИМАЛЬНОГО РАЗМЕРА ПОПУЛЯЦИЙ
закономерно существующее в природе явление, характеризуемое как своеобразный естественный принцип,означающий, что каждому виду животных свойствен специфический для него минимальный размерпопуляции, нарушение которого ставит под угрозу существование популяции, а иногда и вида в целом.
правило популяционного максимума, оно заключается в том, что популяция не может увеличиваться бесконечно, в силу истощения пищевых ресурсов и условий размножения (теория андреварты – бирча ) и ограничения воздействия комплекса абиотических и биотических факторов среды (теория фредерикса).
Вопрос
Итак, как было ясно уже Фибоначчи, прирост популяции пропорционален ее численности, и поэтому, если рост популяции не ограничивают никакие внешние факторы, он непрерывно ускоряется. Опишем этот рост математически.
Прирост популяции пропорционален численности особей в ней, то есть ΔN~N, где N —численность популяции, а ΔN — ее изменение за определенный период времени. Если этот период бесконечно мал, можно написать, что dN/dt=r×N, где dN/dt — изменение численности популяции (прирост), а r— репродуктивный потенциал, переменная, характеризующая способность популяции увеличивать свою численность. Приведенное уравнение называется экспоненциальной модельюроста численности популяции (рис. 4.4.1).
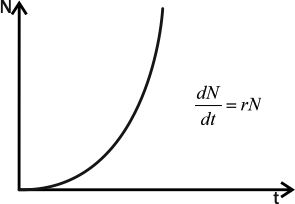
Рис.4.4.1. Экспоненциальный рост.
Как легко понять, с ростом времени численность популяции растет все быстрее, и достаточно скоро устремляется к бесконечности. Естественно, никакое местообитание не выдержит существования популяции с бесконечной численностью. Тем не менее, существует целый ряд процессов популяционного роста, который в определенном временном промежутке может быть описан с помощью экспоненциальной модели. Речь идет о случаях нелимитированного роста, когда какая-то популяция заселяет среду с избытком свободного ресурса: коровы и лошади заселяют пампу, мучные хрущаки — элеватор с зерном, дрожжи — бутыль виноградного сока и т.д.
Естественно, экспоненциальный рост популяции не может быть вечным. Рано или поздно ресурс исчерпается, и рост популяции затормозится. Каким будет это торможение? Практическая экология знает самые разные варианты: и резкий взлет численности с последующим вымиранием популяции, исчерпавшей свои ресурсы, и постепенное торможение прироста по мере приближения к определенному уровню. Проще всего описать медленное торможение. Простейшая описывающая такую динамику модель называется логистической и предложена (для описания роста численности популяции человека) французским математиком Ферхюльстом еще в 1845 году. В 1925 году аналогичная закономерность была заново открыта американским экологом Р. Перлем, который предположил, что она носит всеобщий характер.
В логистической модели вводится переменнаяK — емкость среды, равновесная численность популяции, при которой она потребляет все имеющиеся ресурсы. Прирост в логистической модели описывается уравнением dN/dt=r×N×(K-N)/K(рис. 4.4.2).

Рис. 4.4.2. Логистический рост
Пока Nневелико, на прирост популяции основное влияние оказывает сомножитель r×Nи рост популяции ускоряется. Когда становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K-N)/Kи рост популяции начинает замедляться. Когда N=K, (K-N)/K=0и рост численности популяции прекращается.
При всей своей простоте логистическое уравнение удовлетворительно описывает много наблюдаемых в природе случаев и до сих пор с успехом используется в математической экологии.
15-
№16.Экологическая стратегия выживания—эволюционно сложившийсякомплекс свойств популяции, направленный на повышение вероятности выживания и оставление потомства.
Так А.Г. Раменский (1938) различал три основные типа стратегий выживания среди растений: виоленты, патиенты и эксплеренты.
Виоленты (силовики) — подавляют всех конкурентов, например, деревья, образующие коренные леса.
Патиенты — виды, способные выжить в неблагоприятных условиях («тенелюбивые», «солелюбивые» и т.п.).
Эксплеренты (наполняющие) — виды, способные быстро появляться там, где нарушены коренные сообщества, — на вырубках и гарях (осины), на отмелях и т.д.
Экологические стратегии популяций отличаются большим разнообразием. Но при этом все их многообразие заключено между двумя типами эволюционного отбора, которые обозначаются константами логистического уравнения: r-стратегия и К-стратегия.
| Признак | r-стратегии | K-стратегии |
| Смертность | Не зависит от плотности | Зависит от плотности |
| Конкуренция | Слабая | Острая |
| Продолжительность жизни | Короткая | Долгая |
| Скорость развития | Быстрое | Медленное |
| Сроки размножения | Ранние | Поздние |
| Репродуктивное усиление | Слабое | Большое |
| Тип кривой выживания | Вогнутая | Выпуклая |
| Размер тела | Мелкий | Крупный |
| Характер потомства | Много, мелкие | Мало, крупные |
| Размер популяций | Сильные колебания | Постоянный |
| Предпочитаемая среда | Изменчивая | Постоянная |
| Стадии сукцессии | Ранние | Поздние |