Структура клеточной стенки высших растений
Первоначально клеточная стенка формируется по завершении митоза между двумя клетками и носит название клеточной пластинки. Клеточная пластинка растет центробежно, достигает боковых стенок материнской клетки и сливается с ними; две образовавшиеся дочерние клетки оказываются разделенными. Клеточная пластинка представляет собой единый слой, общий для двух разделяемых ею дочерних клеток. Она образована в основном аморфными пектиновыми веществами, главным образом протопектином и пектатами кальция и магния, имеющими полужидкую консистенцию; целлюлоза в ней отсутствует. Как только края клеточной пластинки достигают продольных стенок, клетки оказываются разделенными и клеточная перегородка называется срединной пластинкой. Она склеивает отдельные клетки в единую ткань и является матриксом будущей клеточной стенки. В начале процесса формирования древесной ткани она состоит, в основном, из пектина, затем, в процессе одревеснения лигнифицируется.
Каждый дочерний протопласт начинает откладывать изнутри свою собственную оболочку, которая примыкает к срединной пластинке. Отложение новых порций материала оболочки происходит, как правило, равномерно по всей поверхности протопласта и такую оболочку растущей клетки называют первичной. Первичная клеточная стенка формируется в процессе роста клеток и обеспечивает поддержание тургора и увеличение растущих клеток в длину. Первичную клеточную стенку и срединную пластинку в древесине часто объединяют под общим названием составной срединной пластины(“compound middle lamella”).
Матрикс первичной клеточной стенки представляет аморфную, пластичную массу, в состав которой входят пектиновые полисахариды, гемицеллюлозы и гликопротеины. Рост клеток происходит растяжением, при этом клеточная стенка растет путем интуссуцепции (явление, когда в пространство между существующими структурами внедряется новый материал). К основным компонентам первичной стенки относятся ксилоглюкан, рамногалактуронан, арабиногалактан, протеогликаны. После окончательного формирования матрикса первичной клеточной стенки в нем появляются микрофибриллы целлюлозы, синтезирующиеся на поверхности протопласта в плазмалемме.
Важным компонентом первичной клеточной стенки является экстенсин – стуктурный гликопротеин, состоящий из ковалентно связанных растворимых предшественников. Последние представляют собой нитевидные молекулы длиной около 80 нм, являющиеся олигомерами декапептида [Ser-(Hyp)4-Val-(Tyr-Lys)2]n (n=30). Углеводная часть состоит из остатков арабинозы, соединенных α-1,2; α-1,3; α-1,4 связями. Тетрасахариды арабинозы присоединены к остаткам оксипролина. Полагают, что при растяжении первичной стенки в процессе роста целлюлозные цепи могут скользить в сетке, которую образует переплетенный с нею и сшитый пероксидазой экстенсин (рис. 3).
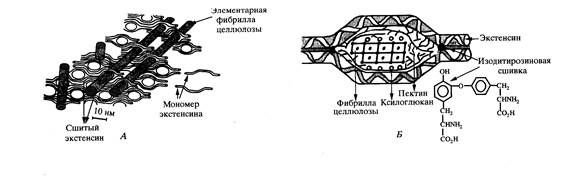
Рис.3. Модель строения первичной клеточной стенки по Лампорту: А- продольное сечение; Б- поперечное сечение элементарной фибриллы целлюлозы, окруженной аморфным полисахаридным матриксом; В- механизм переключения между «слабым» и «сильным» взаимодействием полисахаридного матрикса с сеткой сшитого экстенсина, обеспечивающий подвижность фибриллы при снижении рН (по М.Л. Рабинович и др., 2001).
Связанный с целлюлозой пектин в этих условиях либо фиксирует положение обоих компонентов друг относительно друга через солевые мостики между карбоксилат-анионами уроновых кислот и заряженными аминогруппами остатков лизина белка, либо допускает скольжение при кислом рН, когда солевые мостики размыкаются и образуются димеры карбоксильных групп. Молекулы целлюлозы, погруженные в матрикс первичной клеточной стенки, короткие и состоят примерно из 2000 остатков глюкозы. Микрофибриллы целлюлозы беспорядочно переплетаются между собой, образуя сетчатую структуру.
Первичная клеточная стенка насыщена водой, она легко проницаема для растворенных веществ. В сухом веществе преобладают пектины и гемицеллюлозы, содержание целлюлозы не превышает 30%. Некоторые клетки сохраняют тонкую первичную оболочку до конца жизни, но обычно с возрастом до 70% вещества первичной клеточной стенки замещается лигнином, молекулы которого откладываются в межфибриллярное пространство. Такая лигноцеллюлозная структура является труднонабухающей в воде. После прекращения роста клетки изнутри на первичную клеточную стенку откладываются новые слои веществ, формируя вторичную клеточную стенку (рис.4).
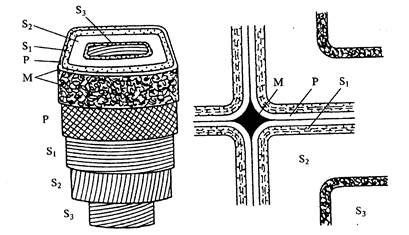
Рис.4. Схема клеточной стенки растений (по Н.А. Родионовой,1989):
М- внутриклеточное вещество срединной пластинки; Р- первичные клеточные стенки; S1-внешний слой вторичной клеточной стенки; S2 – средний слой вторичной клеточной стенки; S3- внутренний слой вторичной клеточной стенки
Для некоторых типов клеток (многие волокна, членики сосудов, трахеиды) образование вторичной оболочки является основной функцией их протопластов, после завершения вторичного утолщения они отмирают. Однако достаточно мощную вторичную оболочку могут иметь и клетки с жизнедеятельным протопластом, такие как паренхимные клетки древесины, флоэмы хвойных растений. Вторичная оболочка может рассматриваться как дополнительная, выполняющая главным образом механическую опорную функцию. В ней содержится значительно меньше воды, чем в первичной оболочке и преобладают микрофибриллы целлюлозы, которые располагаются на близком расстоянии и параллельно друг другу. Содержание целлюлозы во вторичной оболочке составляет обычно 40-50% от массы сухого вещества, гемицеллюлоз – до 20%, лигнина – до 30%, а пектиновых веществ очень мало. В неодревесневших вторичных оболочках (лубяные волокна льна, волоски хлопчатника), содержание целлюлозы может достигать 95%.
Высокое растяжение и строго упорядоченная ориентация микрофибрилл целлюлозы определяет высокие механические свойства вторичных оболочек – прочность на растяжение и эластичность. В лубяных волокнах льна, конопли фибриллы целлюлозы ориентированы параллельно оси клетки, а в стенках механических тканей микрофибриллы ориентированы под углом к оси клетки и образуют спиральную текстуру. В клетках с достаточно развитой вторичной оболочкой, например трахеидах хвойных, можно различить три концентрических слоя, различных по мощности, химическому составу и физическим свойствам (см. рис.3): наружный S1(переходный, относительно тонкий), непосредственно прилегающий к первичной оболочке; средний слой S2 (наиболее мощный); внутренний слой S3 (узкий), граничащий с полостью клетки (люменом). Каждый слой имеет спиральную текстуру.
Средний слой определяет свойства вторичной оболочки. Толщина его, содержание в нем микрофибрилл целлюлозы и лигнина, а так же ориентация микрофибрилл отличаются у разных типов клеток и видов растений, что и обусловливает, например, различия в свойствах древесины у разных пород деревьев. Внутренний слой очень тонок (до 0,1 мкм) и содержит большее количество неструктурированных компонентов, это делает поверхность клеточной стенки, обращенную к люмену, более или менее гладкой. Наиболее высокое содержание целлюлозы у хвойных характерно для внешнего слоя вторичной стенки, а у лиственных - для внутреннего слоя. Высокое содержание полиоз отмечается в слое S1, особенно на границе S1/ S2 .
Оболочки клеток с вторичным утолщением часто подвергаются одревеснению. Особенно характерно оно для клеток, которые после прекращения синтеза оболочки отмирают, например большинство клеток древесины. Лигнификация предохраняет клетки, потерявших тургор от смятия смежными клетками, увеличивает прочность и уменьшает растяжимость. Отложение лигнина происходит обычно с началом вторичного утолщения и наступает, прежде всего, в первичной оболочке, в углах клетки. Затем лигнификация распространяется в срединную пластинку и внутрь (в растущую вторичную оболочку). В конце концов, концентрация лигнина оказывается максимальной в срединной пластинке, постепенно снижаясь по направлению к полости клетки.
Неравномерное утолщение вторичной оболочки приводит к возникновению пор. Те места, куда вещества вторичной оболочки не откладываются, становятся порами. Таким образом, поры – это углубления (впадины, ямки) во вторичной оболочке. У клеток с мощной вторичной оболочкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной оболочки. Если вторичная оболочка в клетке не достигает большой толщины, то поры выглядят как мелкие углубления. По форме порового канала различают поры двух типов – простые и окаймленные. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. Название окаймленной поры происходит оттого, что при рассмотрении ее с поверхности вход в канал поры из полости клетки (внутреннее отверстие) имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде концентрического круга большого диаметра. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, не снижая при этом прочности оболочки.
В теле растения реально существуют хорошо различимые ткани, обладающие целым рядом характерных признаков. Тканями называют устойчивые комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций. Истинные ткани растений подразделяются по функциям на шесть групп: образовательные (меристемы), покровные, проводящие, механические, секреторные (выделительные), базисные (основные).
Образовательные ткани или меристемы состоят из активно метаболизирующих клеток, способных делиться и образовывать новые клетки. С меристемной тканью связан рост растений. Стебли и корни растений обладают верхушечным ростом, листья нарастают благодаря базальному росту, а в стеблях злаков преобладает промежуточный рост. Клетки меристемы тотипотентны (от лат.totum- все, целое, potentium- способность, потенция), то есть они, полностью реализуют свой потенциал развития с образованием целого организма. Культивирование in vitro клеток верхушечной меристемы, свободной от фитопатогенных микроорганизмов в стерильных условиях завершается получением здорового растения, которое используют как посадочный материал. В местах повреждений органов и тканей растений образуются раневые меристемы. На поврежденных участках разрастаются однородные паренхимные клетки, образующие каллус (от лат.callus- мозоль). Каллусные ткани культивируют на питательных средах для получения посадочного и прививочного материала, а так же ценных метаболитов.
Покровные ткани у растений имеют меристемное происхождение. К ним относятся: эпидерма с устьицами, трихомами, эмергенцами на листьях и молодых побегах, перидерма с чечевичками - вторичная покровная ткань на корнях и стеблях преимущественно многолетних растений, корка на ветвях, корнях и стволах многолетних деревьев.
Механические ткани растений представлены колленхимой - уголковой, пластинчатой, рыхлой тканью и склеренхимой. К склеренхиме относятся волокна (например лубяные) и склереиды - структурные элементы механической ткани.
Одними из наиболее важных тканей растений являются проводящие ткани. У растений имеются специализированные ткани – ксилема (от греч.ksilon-дерево), флоэма (от греч.floos –кора, лыко) и проводящие пучки. Проводящие пучки представляют собой обособленные тяжи, состоящие из элементов ксилемы и флоэмы. К флоэме относятся живые (не омертвевшие) ситовидные элементы, паренхимные клетки (лубяная паренхима), сердцевидные лучи и механические элементы. В стеблях флоэма локализуется к наружи от ксилемы. Ксилема состоит из трахеидов – омертвевших клеток, суженных на концах, сосудов - полых трубок и сердцевинных лучей, состоящих из тонкостенных паренхимных клеток.
Трахеида представляет собой сильно вытянутую в длину водопроводящую клетку с ненарушенными первичными стенками. Проникновение растворов из одной трахеиды в другую происходит путем фильтрации через эти стенки, точнее, через окаймленные поры. Сосуд состоит из многих клеток, называемых члениками сосуда. Членики расположены друг над другом, образуя трубочку (рис.5). По сосудам растворы передвигаются значительно легче, чем по трахеидам. В функционирующем состоянии трахеиды состоят лишь из оболочек, так как протопласты распадаются, а полости трахеид заполняются растворами. Трахеиды появились у высших растений в связи с выходом на сушу. Сосуды сформировались значительно позже путем преобразования трахеид. Сосудами обладают почти все покрытосеменные растения. Папаротникообразные и голосеменные растения, как правило, лишены сосудов и имеют только трахеиды.

Рис.5. Анатомические элементы древесины хвойных и лиственных пород :
а – ранняя (весенняя) трахеида; б- поздняя (осенняя) трахеида; в- элементарный сосуд со сложной перфорацией; г- волокно либриформа; д- элементарный сосуд с простой перфорацией; е- паренхимный тяж и паренхимная клетка
Ксилема обеспечивает восходящий ток (от корня к листьям) водных растворов солей, а флоэма – нисходящий ток продуктов фотосинтеза.
Секреторные (выделительные) ткани растений представлены специализированными структурами наружной и внутренней секреции. К числу первых относятся гидатоды, выделяющие водно-солевые растворы; железистые трихомы (железки, нектарники, головчатые волоски), а ко вторым относятся клетки – идиобласты, располагающиеся среди других тканей и накапливающих жидкие или плотные секреты – преимущественно продукты вторичного метаболизма (слизи, оксалат кальция, таннины, терпеноиды, полифенольные соединения); смоляные ходы, эфиромасляные каналы, млечники, пронизывающие все растение.
Базисные ткани растений относятся к разряду мало специализированных. К базисным тканям относят ассимиляционную (хлоренхима), воздухоносную (аэренхима) и запасающие ткани, в которых откладываются белки, жиры, углеводы, пигменты и др.