Особенности анатомического строения древесины
Древесина или вторичная ксилема (от греч. “xylon” – дерево) представляет высокоспециализированную систему растительных тканей. Она состоит из ряда специальных клеток, отличающихся у хвойных (голосеменных) растенийи лиственных (покрытосеменных) деревьев.Древесина и кора иначе флоэма (от греч. “phloios” – кора, лыко) образована клетками камбия–тканью, расположенной между древесиной и корой.
Ежегодно в древесине происходит отложение нового наружного слоя, примыкающего к камбию и убывание возраста слоев от центра к периферии. Древесина хвойных и лиственных пород состоит из годичных концентрических колец, которые хорошо различимы благодаря образованию весенней (ранней) менее плотной и более темной древесины и осенней (поздней) древесины. Ширина годичных колец колеблется от 1 до 10мм и зависит от породы дерева и условий роста: чем лучше условия роста, тем шире годичный слой. Четко выраженные границы годичных колец наиболее характерны для хвойной древесины умеренного пояса. Каждое кольцо имеет плотную и более рыхлую зоны, клетки которых при микроскопическом исследовании существенно отличаются поперечными размерами.
Особенности строения древесины можно наблюдать при распиловке ствола дерева. Для изучения строения древесины используют три среза в трех взаимноперпендикулярных плоскостях: поперечный и два продольных – радиальный (под прямым углом к границе годичных слоев) и тангенциальный (параллельным касательной окружности дерева). Микроскопическое исследование срезов древесины позволяет изучить её анатомические элементы: механические волокна, проводящие элементы, сердцевинные лучи и др. Сердцевинные лучи – радиальные структуры, идущие от центра к периферии, число которых возрастает с ростом диаметра годичного кольца. Они выполняют запасающую функцию: в них, а также в формирующихся перпендикулярно им тяжах осевой паренхимы, характерной для лиственных пород, откладываются запасы питательных веществ, используемых деревом при неблагоприятных условиях.
У большинства хвойных пород на срезах хорошо видны смоляные ходы– вертикальные и горизонтальные каналы с запасами смолы, выделяемой на случай повреждения дерева эпителиальными тканями, примыкающими к смоляным каналам.
Древесина хвойных деревьев имеет сравнительно простое строение и состоит на 90-95% из трахеид(от греч. “tracheios” – горло), мертвых лигнифицированных клеток длиной от 1.5 до 3-4 мм и шириной от 30 до 70 мкм со скошенными торцами и порами в боковых оболочках. В зависимости от сезона трахеиды существенно отличаются толщиной клеточной оболочки и выполняют разные функции. Ранние трахеиды, образующиеся весной и летом, имеют тонкие стенки, широкие полости и являются водопроводящими элементами. Транспорт воды от корней к кроне осуществляется через систему весенних трахеид связанных порами (до 200 на одну весеннюю трахеиду). Простые поры соседних трахеид разделены первичной оболочкой. Окаймленные поры имеют более сложное строение. В центре первичной оболочки у них имеется утолщение – торус(от лат. “torus” – вздутие, выпуклость), а по краям поры – утончение (марго). Торус выполняет функцию одноразового клапана. При необходимости перекрывания потока торус прижимается к выступу вторичной стенки, нависающему над первичной стенкой и диффузия воды прекращается. Вода и низкомолекулярные вещества переносятся через них путем диффузии сквозь марго. Поздние (осенние) трахеиды представляют собой толстостенные образования со значительно меньшим объемом внутренних полостей и меньшим числом пор (от 10 до 50). Они выполняют механическую и запасающую функции.
Живая паренхимная ткань представлена в древесине хвойных преимущественно паренхимой сердцевинных лучей, которая составляет от 4 до 10% общей массы. Эти клетки относительно небольших размеров, с тонкими стенками и большим числом утончений. С окружающими их клетками других типов они сообщаются через полуокаймленные поры, а между собой – через простые поры. В сердцевинных лучах имеются также лучевыетрахеиды, направление оси которых совпадает с направлением лучей. Они обеспечивают транспорт воды в паренхимные ткани.
Строение, форма, число пор на поле перекреста сердцевинных лучей с трахеидами, присутствие или отсутствие в древесине смоляных ходов, однорядное или двурядное рас положение окаймленных пор являются основными диагностическими признаками при определении породного состава древесины хвойных пород. Следует отметить, что диагностические признаки легко установить на радиальном срезе, но лучше изучить все ранее перечисленные три среза (рис.6).
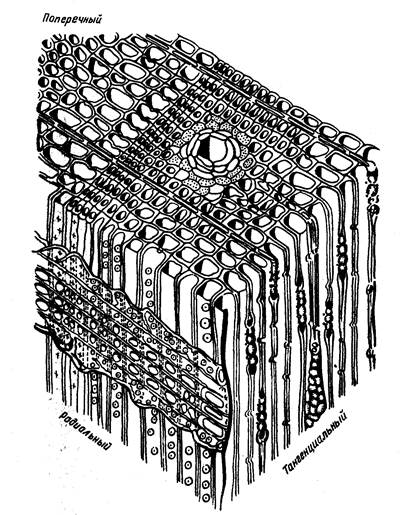
Рис. 6. Схема анатомического строения древесины сосны
Лиственные деревья имеют более твердую древесину при одновременном 5-10-кратном ускорении транспорта воды внутри ствола. В живом лиственном дереве скорость транспорта воды может достигать нескольких десятков метров в час. Древесина лиственных пород по сравнению с древесиной хвойных устроена сложнее (рис.7).
Водопроводящие ткани лиственной древесины состоят из сосудов и сосудистых трахеид, размеры клеток которых подвержены сезонным изменениям. Сосуды образованы путем соединения в продольном направлении широких коротких клеток – члеников сосудов, разделенных перфорированнымии пластинками. Среднее расстояние между перфорированными пластинами, оставшимися от перегородок члеников сосудов варьирует от 10 см (у большинства лиственных) до 3 м (у дуба). Стенки сосудов лигнифицированные, армированы изнутри кольцевидными, спиралевидными или сетчатыми отложениями. Диаметр сосудов у дуба, вяза, ясеня, акации может варьировать от 0,02 (летние или осенние сосуды) до 0,5 мм (весенние сосуды). Такие деревья называются кольцесосудистыми. У большинства лиственных деревьев умеренного климата (береза, клен, ольха, тополь, груша, бук, граб) сосуды мельче и их диаметр относительно мало меняется с сезоном. Эти породы называют диффузнососудистыми или рассеянно-сосудистыми. Древесина, в которой сосуды видны только под микроскопом, называется беспоровой.
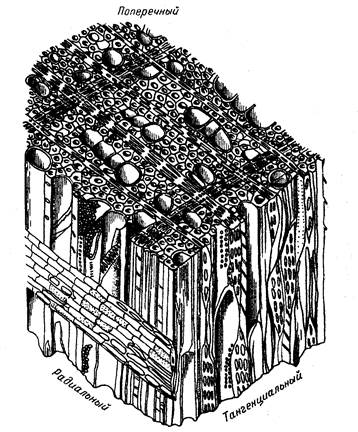
Рис. 7. Схема анатомического строения древесины березы
Механическая прочность лиственной древесины обеспечивается специальными склеренхимными клетками – волокнами либриформа(от лат. “liber” – луб) и волокнистыми трахеидами. Волокна либриформа представляют собой мертвые, вытянутые по длине клетки с заостренными концами и толстыми одревесневшими стенками. Длина волокон либриформа колеблется от 0,3 до 2,6 мм. Либриформ определяет более высокую твердость и плотность лиственной древесины в сравнении с древесиной хвойных пород. Полагают, что либриформ и сосуды лиственных произошли от трахеид голосеменных растений: водопроводящая функция ранних трахеид отошла к сосудам, а механическая функция поздних трахеид – к либриформу.
В лиственной древесине присутствуют волокнистые трахеиды,промежуточные по структуре между трахеидами хвойных и типичными волокнами либриформа. От волокон либриформа они отличаются меньшей толщиной оболочки и наличием мелких окаймленных пор.В древесине березы волокна либриформа и волокнистые трахеиды составляют в сумме от 65 до 70% объема.
Паренхимные клетки в древесине лиственных пород образуют сердцевинные лучи и тяжевую (вертикальную) паренхиму, которая располагается около крупных сосудов и является запасающей тканью. В древесине дуба паренхимная ткань составляет 10-12%, что в 1.5-2 раза больше чем у хвойных деревьев.
В ранней древесине имеются крупные сосуды, которые располагаются кольцом вдоль границы годичного слоя. С возрастом внутренние слои древесины постепенно утрачивают водопроводящую функцию. В старых деревьях ее выполняют только наружные слои более молодой древесины. У ряда пород это приводит к образованию на поперечном срезе четко различимого по цвету ядра (сердцевина), которое окружает живая водопроводящая древесина – заболонь (от англ. “sap” - живица). Созревание древесины, образование ядра у ядровых пород сопровождается закупориванием водопроводящих элементов. Это нормальный физиологический процесс, который заключается в том, что утончения пор, соединяющих сосуды или водопроводящие трахеиды с прилегающими паренхимными клетками, растворяются, и мембраны паренхимных клеток образуют выпячивания внутри проводящей полости, перекрывая ее. Выросты протопластов паренхимной ткани, проникающие через пару пор в полость смежного сосуда, называются тиллами, а сам процесс – затилловыванием древесины. Таким образом, во внутренней спелой, созревшей древесине создаются пробки, препятствующие проникновению воды. У ряда пород тиллы могут заполнять также внутренние полости волокнистых трахеид. В результате резко снижается водопроницаемость древесины. Четко очерченное ядро, иначе спелая древесина, обнаруживается не у всех деревьев: его нет у безъядровых пород.
Основными диагностическими признаками, позволяющими различить древесину лиственных пород являются: строение сосудов, сердцевинных лучей и клетки вертикальной паренхимы. Характерные выросты на внутренней поверхности трахеид сосны и ели, а также волокон либриформа и сосудов бука, выстилающей полость люмена, выделяют в отдельный («бородавчатый») слой. Выросты образуются в период завершения лигнификации клеток и состоят из лигноподобного материала, углеводов и пектиновых веществ. Это подтверждается электронномикроскопическими исследованиями, которые свидетельствуют о четком различии в содержании лигнина между четырьмя основными слоями: составной срединной пластиной, слоем S1, слоем S2 и внутренним слоем, в который включаются и выросты.
Для качественного определения лигнина в растительных тканях используют его цветные реакции, в частности реакцию Мейле, которая позволяет отличить древесину лиственных и хвойных пород. При обработке древесины последовательно 1%-ным раствором перманганата калия, соляной кислотой и раствором аммиака древесина лиственных пород окрашивается в красный цвет, а хвойных – в неопределенно желтый.
Древесина является композиционным материалом и ее прочность обусловлена строением клеточных стенок. Твердым и жестким матриксом, усиливающим прочность клеточных стенок на сжатие и растяжение, является лигнин. Он скрепляет целлюлозные волокна и удерживает их на месте.
Вследствие морфологической неоднородности структур состав и строение древесины различных пород разнятся. В частности, лиственная древесина обычно поддается делигнификации различными методами легче, чем хвойная. Это обусловлено рядом причин: во-первых - мягче древесина, содержание лигнина в лиственной древесине меньше, во-вторых - особенностями строения и локализации лигнина. Лигнин хвойной древесины построен в основном из гваяцилпропановых структурных единиц (рис.5,1) а лигнин лиственных пород – из гваяцилпропановых и сирингилпропановых единиц (рис.8, 1 и 2).

Рис.8. Различия в строении лигнина хвойных и лиственных пород
Большую стойкость структуры лигнина хвойной древесины объясняют также степенью его сшивки. Установлено, что содержание лигнина в срединных пластинках древесины хвойных и лиственных пород примерно одинаково, клеточные же стенки хвойной древесины лигнифицированы в большей степени, чем лиственной.
Материалы с повышенной механической прочностью, подобные материалу клеточной стенки называются композиционными материалами. У таких материалов, состоящих более чем из одного компонента, прочность значительно выше, чем у каждого компонента в отдельности.
Реакционная древесина для повышения устойчивости к механическим нагрузкам включает особые области сжатия, амортизирующие снизу ветви вечнозеленых хвойных пород, и области растяжения, противодействующие провисанию ветвей опадающих деревьев. Более высокую плотность, прочность, повышенное содержание лигнина, пониженное - целлюлозы и, соответственно, более темный цвет имеют области сжатия. Их трахеиды более короткие, округлые по форме, с более толстыми стенками, даже в весенний период. Срединная пластина у них не заполняет полностью межклеточные пространства, оставляя пустоты. Слой S1 тоньше, чем у нормальных клеток, слой S3 отсутствует вовсе, а слой S2 пронизан разветвленными спиральными трещинами, открывающимися в люмен. Области растяжения содержат меньше сосудов, и они мельче, чем в нормальной древесине. Особенностью строения их клеточных стенок является наличие желатиноподобного слоя на внутренней поверхности, обращенной к люмену. Этот слой состоит из ориентированных вдоль оси волокна концентрических ламелл высококристаллической целлюлозы с низким содержанием лигнина и гемицеллюлоз Различные нагрузки сжатие, растяжение сопровождаются разным составом древесины. Так растяжение приводит к снижению доли лигнина, повышению доли целлюлозы, а сжатие повышение содержание лигнина и уменьшению целлюлозы.