Потенциал покоя и потенциал действия
Нервная ткань
А. Структура нервных клеток
Нервная клетка (нейрон) состоит из тела клетки (сомы), отростков (аксонов и дендритов) и концевых пластинок. С помощью дендритов нейроны воспринимают, а посредством аксонов передают возбуждение. На периферии аксоны покрыты шванновскими клетками, образующими миелиновую оболочку с высокими изолирующими свойствами.
Передача возбуждения происходит в нервных окончаниях (синапсах), которые являются местом контакта между нейронами, а также между нейронами и мышечными клетками. В концевых пластинках хранятся химические вещества, нейромедиаторы, выполняющие сигнальные функции. При поступлении нервного импульса медиаторы выделяются в синаптическую щель, передавая возбуждение нейронам или мышечным клеткам.
Для нервных клеток характерно высокое содержание липидов — 50% от сухой массы. Фракция липидов включает разнообразные фосфо-, глико- и сфинголипиды.
Б. Энергетический обмен головного мозга
Головной мозг хорошо снабжается кровью и имеет интенсивный энергетический обмен. Хотя головной мозг составляет около 2% массы тела, при спокойном состоянии организма он утилизирует около 20% поглощенного кислорода и 60% глюкозы, которая полностью окисляется до СО2 и Н2О в цитратном цикле и путем гликолиза.
В клетках головного мозга практически единственным источником энергии, который должен поступать постоянно, является глюкоза. Только при продолжительном голодании клетки начинают использовать дополнительный источник энергии — кетоновые тела (см. рис. 305). Запасы гликогена в клетках головного мозга незначительны. Жирные кислоты, которые в плазме крови транспортируются в виде комплекса с альбумином, не достигают клеток головного мозга из-за гематоэнцефалического барьера. Аминокислоты не могут служить источником энергии для синтеза АТФ (АТР), поскольку в нейронах отсутствует глюконеогенез. Зависимость головного мозга от глюкозы означает, что резкое падение уровня глюкозы в крови, например, в случае передозировки инсулина у диабетиков, может стать опасным для жизни.
В клетках центральной нервной системы наиболее энергоемким процессом, потребляющим до 40% производимого АТФ, является функционирование транспортной Na+/К+-АТФ-азы (Na+/K+-«насоса») клеточных мембран. Активный транспорт ионов Na+ и К+ компенсирует постоянный поток ионов через ионные каналы. Кроме того, АТФ используется во многих биосинтетических реакциях.
В. Метаболизм аминокислот
В клетках головного мозга идет активный метаболизм аминокислот. В головном мозге концентрация аминокислот почти в 8 раз выше, чем в плазме крови, и существенно выше, чем в печени. В особенности высоким является уровень глутамата (примерно 5-10 мМ) и аспартата (2-3 мМ). Эти аминокислоты образуются в реакции трансаминирования из промежуточных метаболитов цитратного цикла, 2-оксоглутарата и оксалоацетата.
В тканях мозга интенсивно протекают метаболические превращения аминокислот, такие, как окислительное дезаминирование, трансаминирование, модификация боковой цепи и др. В особенности важной для нормального функционирования головного мозга является реакция декарбоксилирования, в результате которой образуется γ-аминомасляная кислота (γ-аминобутират) (ГАМК, GABA) (предшественник — глутамат) и биогенные амины. Биосинтез и деградацию глутамата можно рассматривать, как побочный путь цитратного цикла (ГАМК-шунт), который в отличие от основного цикла не приводит к синтезу гуанозин-5'-трифосфата (см. рис. 139). ГАМК-шунт характерен для клеток центральной нервной системы, но не играет существенной роли в других тканях.
Некоторые аминокислоты, например глицин, аспартат, глутамат, ГАМК, выполняют в нейронах функцию медиаторов. Они хранятся в синапсах и выделяются при поступлении нервного импульса (см. рис. 343). Переносчики индуцируют или ингибируют потенциал действия, контролируя тем самым возбуждение соседних нейронов.
Потенциал покоя и потенциал действия
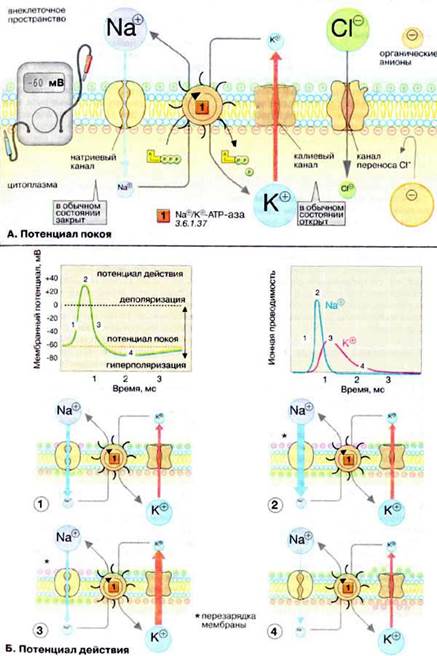
А. Потенциал покоя
Мембраны, в том чикле плазматические, в принципе непроницаемы для заряженных частиц. Правда, в мембране имеется Na+/K+-АТФ-аза (Nа+/К+-АТР-аза), осуществляющая активный перенос ионов Na+ из клетки в обмен на ионы К+. Этот транспорт энергозависим и сопряжен с гидролизом АТФ (АТР) (см. рис. 221). За счет работы «Nа+,К+-насоса» поддерживается неравновесное распределение ионов Na+ и К+ между клеткой и окружающей средой (см. с. 128). Поскольку расщепление одной молекулы АТФ обеспечивает перенос трех ионов Na+ (из клетки) и двух ионов К+ (в клетку), этот транспорт электрогенен, т. е. цитоплазма клетки заряжена отрицательно по отношению к внеклеточному пространству.
Электрохимический потенциал. Содержимое клетки заряжено отрицательно по отношению к внеклеточному пространству. Основная причина возникновения на мембране электрического потенциала (мембранного потенциала Δψ) — существование специфических ионных каналов. Транспорт ионов через каналы происходит по градиенту концентрации или под действием мембранного потенциала. В невозбужденной клетке часть К+-каналов находится в открытом состоянии и ионы К+ постоянно диффундируют из нейрона в окружающую среду (по градиенту концентрации). Покидая клетку, ионы К+ уносят положительный заряд, что создает потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости различных ионов (см. таблицу на рис. 129) видно, что каналы, проницаемые для Na+ и Cl-, преимущественно закрыты. Ионы фосфата и органические анионы, например белки, практически не могут проходить через мембраны. С помощью уравнения Нернста (см. рис. 129) можно показать, что мембранный потенциал нервной клетки в первую очередь определяется ионами К+, которые вносят основной вклад в проводимость мембраны.

Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na+, К+, Са2+ и Cl-. Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (или электровозбудимые), например быстрые Na+-каналы, и лиганд-управляемые (или хемовозбудимые), например никотиновые холинэргические рецепторы. Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами (см. рис. 343), белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — «закрыт» — и т.д.).
Б. Потенциал действия
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия. Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа+-канала (1). Ионы Na+ устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала (2). При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время (3). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы (2) и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциал принимает первоначальное значение (3), и даже превышает на короткое время потенциал покоя (4). После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Медиаторы нервной системы