Представники: міноги, міксини
Клас Головохордові
Представник: ланцетник
Нервова система ланцетника представлена головним мозком, що є розширеним переднім кінцем нервової трубки, та спинним мозком. У ланцетника – структура, що містить нейросекреторні елементи, є гомологічною ГГНС хребетних. Вона носить назву «інфундибулярний орган», який розташований навколо заглиблення вентральної стінки розширеної порожнини переднього кінця нервової трубки. «Інфундибулярний орган» складається з великих епендимоподібних клітин, що своєю апікальною частиною досягають порожнини шлуночка, а їхня базальна частина переходить у відросток - аксон, що йде у вентральному напрямку до поверхні нервової трубки. В цитоплазмі цих клітин виявляються дрібні пилоподібні гоморіпозитивні гранули, що розглядаються як секреторні включення .
Підтип Черепні (Craniata)
Клас Круглороті (Cyclostomata)
Представники: міноги, міксини
У круглоротих, як і у всіх нижчих тварин у гіпоталамусі є парне крупноклітинне преоптичне ядро - ПЯ, що розташоване в бічних стінках передньої частини проміжного мозку трохи ростро-дорсальніше перехрестя зорових нервів у безпосередній близькості від порожнини III шлуночка мозку. У ділянці ядра виявлена слабка васкуляризація. Поряд з великими нейросекреторними клітинами, розташованими переважно в дорсо-каудальній частині, є багато більш дрібних нейроцитів, зосереджених поблизу преоптичної бухти III шлуночка. Межі самого ядра, виражені не дуже чітко.
Нейросекреторні клітини переважно біполярні. Їхні довгі відростки – аксони, як правило, відходять вентро-латеральному напрямку убік зорових трактів. Основна маса коротких відростків – дендритів орієнтована у медіальному напрямку, убік порожнини Ш шлуночка. Така форма контакту називається дендро-вентрикулярними контактактами. Досить часто НСК розташовано безпосередньо серед клітин епендими.
У круглоротих - нейросекреторні включення мають вигляд дрібних гранул і великих крапель.
Аксони» НСК преоптичного ядра, обігнувши оптичні тракти формують преоптико-гіпофізарний тракт (преоптико-інфудибулярний по Bargmann, 1953), що потім направляється каудально до гіпофіза. Частина аксонів у постоптичній ділянці вступає в контакт із капілярами оболонки мозку, утворючі аксо-вазальні контакти. Ці капіляри оболонки мозку зв'язані з передньою часткою гіпофіза, тобто є гомологами портальної системи аденогіпофіза інших хребетних. Сама ж зона, у якій локалізовані контактуючі з капілярами нейросекреторні закінчення позначається як проксимальною нейросекреторною контактною ділянкою,що є гомологічною середин ному підвищенню амфібій та Amniota.
За даними окремих дослідників окрім ПЯ, у міног у каудальному відділі гіпоталамуса є невелика група НСК заднього гіпоталамічного ядра(Nucleus posteior hypothalamicus). Дендрити цих клітин підходять до III шлуночка, тоді як аксони направляються в нейрогіпофізарний шар та до вентральної поверхні мозку.
Крім того, від преоптичного ядра відходять нейросекреторні волокна, що утворять екстра-гіпоталамічні нейросекреторні тракти:
- гіпоталамо-теленцефальний тракт (до гіпокампу переднього моз ку);
- гіпоталамо-хабенулярний тракт (у хабенулярний ганглій);
- гіпоталамо-мезенцефальний тракт (до оптичної покришки);
- гіпоталамічний тракт у довгастий мозок .
У круглоротих практично немає сформованого нейрогіпофіза, але існує диференційований аденогіпофіз, що має епітеліальне походження.
Аденогіпофіз є досить компактним комплексом клітинних острівців і трабекул, оточеним сполучнотканинною капсулою. У дорослих форм круглоротих досить чітко виявляється диференціація аденогіпофіза на три відділи: про-, мез-, метааденогіпофіз Проаденогіпофіз та мезаденогіпофізвідповідає передній частці, метаденогіпофіз – проміжній частці. Артерії входять у щілину між двома основними компонентами гіпофіза і розпадаються на мережу дрібних судин, що посилають окремі гілочки у вентральному напрямку до епітеліальних залозистих клітин. Отже, фізіологічна взаємодія між основними відділами гіпофіза можливо тільки через мережу кровоносних судин.
Нейрогіпофізу відповідає пластинкоподібний шар нервових волокон, що лежить між епендимой, що вистилає вентральну стінку воронки, і дорсальною поверхнею аденогіпофіза. Цей шар називаємо нейрогіпофізарним. Він єдистальною нейросекреторною контактною ділянкою. В ньому зосереджені нейросекреторні волокна (аксони) і їхні закінчення, що контактують з численними синусоїдними капілярами (аксо-вазальні контакти), розташованими на межі проміжної частки гіпофіза (відповідає метааденогіпофізу по Пікфорд, 1957). До того ж контактна поверхня не завжди рівна, тому що місцями нейрогіпофізарна тканина неглибоко вдасться й епітеліальний шар метаденогіпофіза.
Таким чином, вже в представників Anamnia (нижчі хордові) і зокрема у безчерепних виявлені клітинні елементи, секреторна функція і топографія яких, свідчать про їхню гомологію з нейроцитами ПЯ чи супраоптичними (СОЯ) і паравентрикулярними (ПВЯ) ядрами гіпоталамуса вищих хребетних.
Клас Риби (Pisces)
Будова ГГНС різних риб має загальні риси, але все-таки істотно розрізняється в представників великих таксономічних груп.
Пластинчастозяброві (Elasmobranchii). В акул (Scyliorhinus sttellaris, Mustelus mustelus) і скатів (Raja clavata, Trygon pastinaca) НСК гіпоталамуса зосереджені в парному ПЯ.Волокна збираються в преоптико-гіпофізарний тракт, що проходить у каудальному напрямку в стінці дна III шлуночка поблизу поверхні мозку. У них існує типоваа проксимальна контактна ділянка і примітивна система портальних судин аденогіпофіза.
Аденогіпофізмає порожнину, тобто між про- та мезоаденогіпофізом (дистальна частка) та метаденогіпофізом (проміжна частка) існує щілина. Епітеліальні тяжі аденогіпофіза перетворюються на типові фолікули.
Нейрогіпофіз має вигляд пластинкоподібного виросту, що відповідає дистальній нейросекреторній контактній ділянці.
Хрящові (Chondrosteoidei). У представників осетрових (осетер, севрюга, білуга) нейросекреторним центром є ПЯ. Однак у каудальній частині гіпоталамуса поблизу воронки існує невелика парна група клітин- мамілоінфундибулярне ядро (Nucleus mammilloinfundibularis). У вентро-ростральній частині сірого бугра формується преоптико-гіпофізарний тракт,який в каудальному напрямку заходить в нейрогіпофіз.
В осетрових є проксимальна контактна ділянкарозвинена краще,вона значно ширше і поширюється каудальне, ніж у .пластинчастозябрових риб. Тут значно більше НСК-терміналей, що контактують з капілярами.
Більшість осетрових риб мають аденогіпофіз з порожниноюаналогічний гіпофізу пластинчатозябрових. Новою особливістю для осетрових риб є взаємне проростання епітеліальних тяжів у тканину нейрогіпофіза.
За будовою нейрогіпофізпредставлений дуже масивними трубчастими структурами, що занурюються у проміжну частку.
Костисті (Teleostei). Особливе місце серед риб займають костисті, котрі за своїм положенням у філогенетичній системі найбільш віддалені від основного стовбура еволюції хребетних.
У гіпоталамусі є добре розвинене парне ПЯ,в якому виявляється вентральна – дрібноклітинна частина(складається з дрібних НСК) та дорсальна – крупноклітинна частина(складається з крупних НСК). Однак у багатьох видів є ще друге дуже крупне непарне нейросекреторне ядро – латеральне ядро сірого бугра (Nucleus lateralis tuberis). У цьому ядрі, також як і в преоптичному, спостерігається полярність у розташуванні його клітинних елементів.
При цьому відзначають інтенсивну васкуляризацію ядер, що проявляється у тісному контакті капілярів з тілами клітин.
Основна маса аксонів НСК формує парні преоптико-гіпофізарні тракти. Ці тракти проходять через латеральне ядро сірого бугра і тільки у вентральній частині останнього возз'єднуються з туберо-гіпофізарним трактом, що вперше з’являється у костистих риб. Нервові волокна цього тракту прямують також до нейрогіпофіза, але деяка їх частина закінчуються на капілярах, які направляються до про- і частково до мезоаденогіпофіза.
Аденогіпофіз,як і у попередніх видів риб,має три відділи: про-, мезо- та метааденогіпофіз.
Нейрогіпофіз у різних видів костистих має дуже різноманітну будову від древоподібного до компактного утворення. Від нейрогіпофіза вглиб залозистої паренхіми аденогіпофіза відходять найтонші розгалужені термінальні гілочки.
Функцію портальної системи в костистих риб виконує добре розвинене венозне сплетення, загальне для нейрогіпофіза і залозистих елементів метааденогіпофіза та мезоаденогіпофіза. Проаденогіпофіз постачається гілочками внутрішньої сонної артерії, що потім впадають у вищевказане венозне сплетення. Венозна кров з нейро- і аденогіпофіза збирається в гіпокраніальну вену і далі, зрештою, попадає в загальну циркуляцію. Отже можна відзначити особливе кровопостачання нейрогіпофіза та аденогіпофіза.
У деяких костистих описані окремі нейросекреторні екстрагіпоталамічні тракти, що йдуть від крупно клітинної чатсини ПЯ до субкомісурального органу і хабенулярному ганглію.
Таким чином, в процесі філогенеза риб має місце ослаблення відтоку нейросекрета через епендиму в III шлуночок мозку і посилення транс портування гіпоталамічних гормонів по асконам у гіпофіз. Вперше у костис тих риб формується нейрогіпофіз та туберо-гіпофізарний тракт, гомолог портальної системи аденогіпофіза.
Клас Амфібії
У всіх земноводних є парне масивне ПЯ (преоптичне ядро). Його топографія загалом подібна з топографією в риб. Однак в амфібій НСК, особливо в дорсальній частині, пухко розташовані. Намічається відокремлення двох груп клітин – дорсальної і вентральної, з яких згодом у Amniota формуються СОЯ та ПВЯ.
Основна маса аксонів НСК ПЯ поблизу серединного підвищення формують преоптико-гіпофізарний тракт, який має вигляд суцільного шару, розділеного відростками клітин епендими на окремі пучки.
Окремі відростки нейронів ПЯ в земноводних також утворять екстрагіпоталамічні тракти:
1) гіпоталамо-гіпокампальний до комісури гіпокампа;
2) гіпоталамо-септальний до субфорнікального органу;
3) гіпоталамо-хабенулярний до хабенулярного ядра;
4) гіпоталамо-теленцефальний у передній мозок;
5) гіпоталамо-диенцефальні асоціативні тракти.
Більшість нейросекреторних волокон цих трактів закінчуються або на капілярах, або поблизу епендими, в основному III і, рідше, бічних шлуночків, утворюючі аксо-вазальні чи аксо-вентрикулярні контакти. В амфібій досить добре виражені дендро-вентрикулярні контакти.
В амфібій, вперше в ряді хребетних, формується компактний нейрогіпофіз, у якому чітко виражені три частки, характерні для вищих хребетних: серединне підвищення, інфундибулярна частина і головна задня частина.На відміну від риб в ембріогенезі амфібій закладка нейрогіпофіза є частиною загального анатомічного цілого з гіпоталамусом.
У масивному серединному підвищенні: чітко розрізняються три шари: 1) шар епендими, що виступає інфундибулярну бухту;
2) шар нейросекреторних,волокон преоптико-гіпофізарного тракту;
3) зовнішній нейроваскулярний шар.
В зовнішньому шарі СП вперше з’яляється примітивнапортальна система аденогіпофіза.Тут є значна кількість аксо-вазальних контактів із синусоїдами первинного капілярного сплетення, що дають початок 2-3 портальним венам передньої частки аденогіпофіза.
Отже, доведено наявність у амфібій двох незалежних джерел кровопостачання нейрогіпофіза та проміжної частки аденогіпофіза й окремо серединного підвищення.
У складі аденогіпофізаєтуберальна, дистальна та проміжна частки.
У дорослих жаб при застосуванні різноманітних забарвлень виявлено шість типів клітин.
Диференціювання базофілів і оксифілів посилюється перед метаморфозом. З настанням метаморфоза, тобто в період інтенсивного перетворення ї водяної форми в сухопутну, у багатьох нейронах ПЯ збільшується кількість гранул і крапель нейросекреторної субстанції. Нейрогіпофіз, особливо його задня частка, містить багато гранульованого і дифузно розташованого нейросекрету.
Таким чином, вперше у амфібій формується компактний нейрогіпофіз та СП. В зовнішньому шарі СП вперше з’яляється примітивна портальна система аденогіпофіза.
ВИЩІ ХРЕБЕТНІ (AMNIOTA)
Клас Плазуни (Reptilia)
У рептилій вперше серед хребетних з'являються два нейросекреторних центри, характерних для усіх (Amniota): парні СОЯ і ПВЯ, що є гомологами ПЯ гіпоталамуса Anamnia.
Однак між обома ядрами розташовані як окремі НСК, так і невеликі групи їх, що як би вказують на походження цих нейроцитів з єдиного ПЯ Anamni.
Супраоптичне ядро представлене двома парними скупченнями крупних НСК, які симетрично розташовані безпосередньо над оптичною хіазмою.
Паравентрикулярне ядро розташоване у бічних стінках III шлуночка, безпосередньо під його епендимою, займаючи більш дорсальне положення порівнянно із СОЯ, тобто вище над ним.
Обидва ядра, і особливо СОЯ, на відміну від інших ділянок гіпоталамуса є компактними, а також багато васкуляризовані. Клітини обох ядер мають подібні за будовою. Їхні розміри незначно варіюють. Нейросекрет представлений гоморіполозитивними гранулами і, рідше, великими краплями.
Переважна маса аксонів СОЯ та ПВЯ формують гіпоталамо-гіпофізарний тракт (супраоптикопаравентрикуло-гіпофізарний тракт), а менша частина, екстрагіпоталамічні тракти.
У рептилій описані наступні екстрагіпоталамічні тракти, що беруть початок головним чином від ПВЯ:
1) гіпоталамо-парафізарний (до парафізу);
2) гіпоталамо-септальний (до субфорнікального органу);
3) гіпоталамо-хабенулярний;
4) гіпоталамо-теленцефальний, волокна якого проходячи уздовж стінок бічних шлуночків, доходять майже до нюхових часток.
Крім того, у представників цього класу з'являються нові тракти, що йдуть за межі гіпоталамуса в каудальному напрямку до стовбура мозку.
Закінчення нейросекреторних волокон утворюють аксо-вентрикулярні й аксо-вазальні контакти. Дуже рідко волокна закінчуються на тілах нервових клітин. Такий тип контакту називають аксо-соматичний нейросекреторний контакт.
СП та інфундибулярна частина нейрогіпофіза в рептилій, як і в амфібій, мають тришарову будову. Однак у рептилій ці два відділи нейрогіпофіза, як правило, мають більшу площу, що пояснюється значною роллю портальної системи в представників цього класу хребетних.
Ділянка СП по морфологічній відособленості від первинної капілярного сплетення близька до аналогічної організації у птахів та істотно відрізняється від амфібій і ссавців.
Зовнішній шар СП контактує з невеликою епітеліальною пластинкою аденогіпофіза, що включає первинне сплетення кровоносних судин, що йдуть від каротидної артерії. Від первинного сплетення відходить пучок портальних вен, які направляються в дистальний відділ аденогіпофіза. Дистальна частка аденогіпофіза і нейрогіпофіз не контактують між собою, будучи відділені невеликою щілиною. Каудальний відділ дистальної частки, в дорсальному напрямку, переходить безпосередньо в паренхіму проміжної частки.
Своєрідність мікроскопічної структури аденогіпофіза полягає в тім, що він утворений безліччю досить тонких тяжів-трабекул, роз'єднаних кровоносними судинами. У таких тяжах, поряд з типовими хромофобами, розташовані невеликі за розміром секреторні клітини (базофіли й ацидофіли), які знаходяться по периферії тяжів.
Особливістю аденогіпофіза рептилій також є в будові проміжною частки. Проміжна частка на фронтальних зрізах має вигляд чашеподібного футляру, що охоплює нейрогіпофіз.
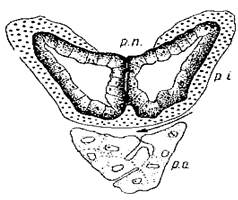
Рис. Будова гіпофіза рептилій (фронтальний зріз).
Відомо, що в більшості птахів проміжна частка гіпофіза взагалі відсутня. У рептилій же вона розвинена більше, ніж у ссавців й амфібій.
Таким чином, організація нейросекреторної системи у плазунів значно відрізняється від такої в риб і земноводних. Вона має багато загальних морфологічних рис з нейросекреторною системою птахів. У рептилій вперше з'являються характерні для усіх (Amniota) парні СОЯ і ПВЯ. Виділяють до 8 ядер. Гіпофіз має добре розвинену проміжну частку. СП та портальна система набувають подальшого розвитку.
Клас Птахи (Aves)
Дослідження на птахах показали, щовони володіють добре розвинутою ГГНС, що представлена крупно- та дрібноклітинними НСК.
Крупноклітинні ядра: СОЯ та ПВЯ гіпоталамуса.НСК цих ядер держать досить пухко та між ними існують перехідні численні нейросекреторні клітини. Більшість НСК біполярні.
СОЯ розташовано дорсальніше оптичної хіазми, більш розвинене, тут зустрічаються мультиполярні нейроцити. НСК СОЯ витягнуті в горизонтальному напрямку вздовж оптичного тракту.
ПВЯорієнтовані вертикально і паралельно стінки III шлуночка мозку. У його складі виділяють декілька груп.
Хоча у СОЯ також прийнято виділяти медіальну та латеральну групи.
Функції:крупноклітинні ядравідіграють роль в регуляції сезонних біологічних явищ: у період розмноження більш активується СОЯ, тоді як у період линяння – ПВЯ, у птахів мігруючих – обидва ядра. Відомо, що тут виробляються нонапептиди: аргінін-вазотоцин і мезотоцин.
Аксони НСК обох ядер беруть участь в формуванні супраоптикопаравентрикуло-гіпофізарного тракту, волокна якого прямують до нейрогіпофіза.
Дрібноклітинні ядраможна поділити на дві групи, що розташовані у ділянці сірого бугра та ядра переднього гіпоталамуса.
Першу групу складають:аркуатне (туберальне або інфундибулярне) ядро – АЯ; вентромедіальне (ВМЯ); дорзомедіальне ядро – ДМЯ.
Другу групу- супрахіазматичне ядро - СХА; медіальне, латеральне та перивентрикулярне преоптичне ядро.
Серед цих НСК є моноамінергічні клітини.
Аксони дрібноклітинних ядер утворюють туберо-інфундибулярний тракт, що забезпечує транспорт рилізінг-гормонів гіпоталамуса до портальної системи аденогіпофіза у СП.
Функції:НСК дрібноклітинних ядер вироблють ліберини та статини та беруть участь в регуляції вегетативних функцій, сезонних ритмів, розмноження та росту.
У птахів виявлені екстрагіпоталамічні тракти:
1) гіпоталамо-септальний (у субфорнікальний орган);
2) гіпоталамо-теленцефальний (;
3) гіпоталамо-хабенулярний.
Значного розвитку порівняно з іншими хребетними досягає СП. У ньому розрізняють два відділи: передній та задній. За будовою в СП розрізняють три чи п’ять шарів: епендимний (вистиляє порожнину III шлуночка); внутрішній (тут проходять транзитом волокна СОПВ-гіпофізарного тракту); зовнішній (палісадний).Додатково виділяютьрети кулярний, фіброзний.
У птахів на відміну від ссавців капіляри первинного портального сплетення лежать на поверхні СП, тобто не занурюються у паренхіму.
До того ж портальна система набуває подальшого розвитку, тобто чітко розрізняють первинне сплетення капілярів, портальні вени та вторинне сплетення капілярів.
Аденогіпофіз.У його складі добре розвиненою є дистальна частка, де знаходяться різні аденотропоцити, які синтезують тропні гормони. Проміжна частка відсутня. Функція аденогіпофіза регулюється гіпоталамічними гормонами.
Нейрогіпофіз. Має вигляд компактної дещо сплющеної в дорсовентральному напрямку структури. Нейрогіпофіз повністю відокремлений від аденогіпофіза.