СТРОЕНИЕ , СВОЙСТВА И ФУНКЦИИ
Углеводы или сахариды - одна из основных групп органических соединений организма. Они представляют собой первичные продукты фотосинтеза и исходные продукты биосинтеза других веществ в растениях ( органические кислоты, аминокислоты), а также содержатся в клетках всех других живых организмов. В животной клетке содержание углеводов колеблется в пределах 1-2%, в растительной оно может достигать в некоторых случаях 85-90% массы сухого вещества.
Углеводы состоят из углерода, водорода и кислорода, причем у большинства углеводов водород и кислород содержатся в том же соотношении, что и в воде ( отсюда их название - углеводы ). Таковы, например, глюкоза С6Н12О6 или сахароза С12Н22О11. В состав производных углеводов могут входить и другие элементы. Все углеводы делятся на простые (моносахариды) и сложные (полисахариды).
Среди моносахаридов по числу углеродных атомов различают триозы (3С), тетрозы (4С), пентозы (5С), гексозы (6С) и гептозы (7С). Моносахариды с пятью и более атомами углерода, растворяясь в воде, могут приобретать кольцевую структуру. В природе наиболее часто встречаются пентозы ( рибоза, дезоксирибоза, рибулоза) и гексозы ( глюкоза, фруктоза, галактоза). Рибоза и дезоксирибоза играют важную роль в качестве составных частей нуклеиновых кислот и АТФ. Глюкоза в клетке служит универсальным источником энергии. С превращением моносахаридов связаны не только обеспечение клетки энергией, но и биосинтез многих других органических веществ, а также обезвреживание и выведение из организма ядовитых веществ, проникающих извне или образующихся в процессе обмена веществ, например, при распаде белков.
Ди- и полисахариды образуются путем соединения двух и более моносахаридов, таких, как глюкоза галактоза маноза, арабиноза или ксилоза. Так, соединяясь между собой с выделением молекулы воды, две молекулы моносахаридов образуют молекулу дисахарида. Типичными представителями этой группы веществ являются сахароза ( тростниковый сахар), мальтаза (солодовый сахар), лактоза (молочный сахар). Дисахариды по своим свойствам близки к моносахаридам. Например, и те, и другие хорошо растворимы в воде и имеют сладкий вкус. К числу полисахаридов принадлежит крахмал, гликоген, целлюлоза, хитин, каллоза и др.
Основная роль углеводов связана с их энергетической функцией. При их ферментативном расщеплении и окислении выделяется энергия, которая используется клеткой. Полисахариды играют главным образом роль запасных продуктов и легко мобилизуемых источников энергии ( например, крахмал и гликоген), а также используются в качестве строительного материала ( целлюлоза, хитин). Полисахариды удобны в качестве запасных веществ по ряду причин: будучи нерастворимы в воде, они не оказывают на клетку ни осмотического, ни химического влияния, что весьма важно при длительном хранении их в живой клетке: твердое, обезвоженное состояние полисахаридов увеличивает полезную массу продуктов запаса за счет экономии их объема. При этом существенно уменьшается вероятность потребления этих продуктов болезнетворными бактериями и другими микроорганизмами, которые, как известно, не могут заглатывать пищу, а всасывают вещества всей поверхностью тела. И наконец, при необходимости запасные полисахариды легко могут быть превращены в простые сахара путем гидролиза.
Классификация: Моносахариды:глюкоза,фруктоза,галактоза,Дисахариды:сахароза,лактоза.мальтоза, Полисахариды:крахмал.гликоген,пектиновые вещества,клетчатка.
2….. биологическое значение углеводов :
1 Углеводы выполняют пластическую функцию , то есть участвуют в построении костей, клеток, ферментов. Они составляют 2-3 % от веса.
2 Углеводы являются основным энергетическим материалом. При окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.
3В крови содержится 100-110 мг глюкозы. От концентрации глюкозы зависит осмотическое давление крови.
4Пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ.
5 Углеводы выполняют защитную роль в растениях.
3……..Всасывание углеводов пищи в желудочно-кишечном тракте - это транспорт углеводов из полости желудочно-кишечного тракта во внеклеточную (межклеточную жидкость, кровь и лимфу) и внутриклеточную жидкости организма. Всасывание углеводов завершает метаболизм углеводов в системе пищеварения.
Метаболизм углеводов - это совокупность ферментативных реакций, протекающих в организме, исходным метаболитом (объектом изменения) которых являются углеводы.
Начальным этапом метаболизма углеводов является метаболизм углеводов в системе пищеварения. Это совокупность процессов переваривания полисахаридов и олигосахаридов пищевых продуктов, поступающих в пищеварительный тракт с пищей, до моносахаридов и их последующее всасывание из полости пищеварительного тракта в кровь и лимфу. Метаболизм углеводов в системе пищеварения сопряжён с метаболизмом углеводов в тканях, клетках и субклеточных структурах (митохондрии, цитозоль, эндоплазматический ретикулум и др).
Переваривание углеводов пищи в желудочно-кишечном тракте - это часть метаболизма углеводов , совокупность управляемых процессов, осуществляемых в системе пищеварения, которые представляют собой химическую переработку углеводов , поступающих в организм с пищей, для последующего их всасывания в кровь и в лимфу.
Пищевые вещества содержат крупномолекулярные белки, углеводы и липиды, которые не способны к всасыванию в кровь и лимфу из-за больших размеров своих молекул. Химическая переработка пищи в желудочно-кишечном тракте представляет собой последовательное ступенчатое ферментативное гидролитическое расщепление крупномолекулярных белков, углеводов и липидов до простых веществ, способных к всасыванию.
В числе углеводов потребляемой пищи содержатся полисахариды крахмал и гликоген. Расщепление этих углеводов начинается в полости рта и продолжается в желудке. Катализатором гидролиза является фермент амилаза слюны. При расщеплении из крахмала и гликогена образуются декстрины и в небольшом количестве - мальтоза. Пережеванная и смешанная со слюной пища проглатывается и попадает в желудок. Проглоченные пищевые массы со стороны поверхности полости желудка постепенно перемешиваются с желудочным соком, содержащим соляную кислоту. Содержимое желудка с периферии приобретает значительную кислотность (pH = 1,5 ÷ 2,5). Такая кислотность дезактивирует амилазу слюны. В то же время в толще массы желудочного содержимого амилаза слюны некоторое время продолжает действовать и происходит расщепление полисахаридов с образованием декстринов и мальтозы. Желудочный сок не содержит ферментов, расщепляющих сложные углеводы. Поэтому гидролиз углеводов с увеличением кислотности в желудке прерывается и возобновляется в двенадцатиперстной кишке.
Для расщепления каждой группы веществ существуют свои определенные ферменты : для белков – протеазы, для жиров – липазы, для углеводов – карбогидразы. Основными железами, которые вырабатывают пищеварительные ферменты , являются железы ротовой полости (слюнные железы), железы желудка и тонкого кишечника, поджелудочная железа и печень. Главную роль в этом играет поджелудочная железа, которая вырабатывает не только пищеварительные ферменты , но и гормоны, такие, как инсулин и глюкагон, участвующие в регуляции белкового, углеводного и липидного обменов.
Клеток, вырабатывающих пищеварительные ферменты в поджелудочной железе, достаточно много. Они образуют специальные скопления, от которых отходят маленькие выводные протоки; по ним передвигается секретируемый сок поджелудочной железы, представляющий собой своеобразный коктейль из разных ферментов .
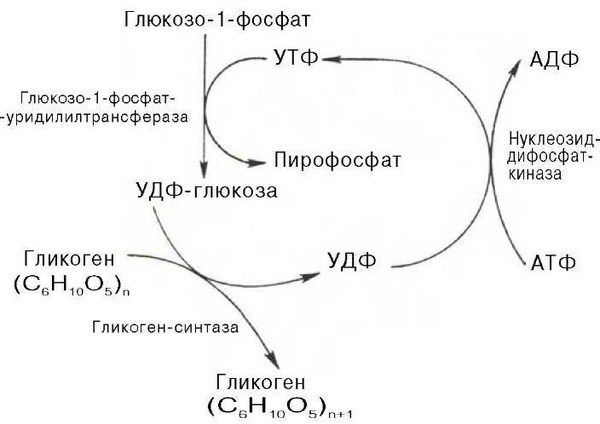
55555 
7777.. Поскольку одним из главных видов биологического «топлива» служат углеводы , мы рассмотрим биологическое окисление на примере того, как окисляется в клетках глюкоза и как запасается при этом энергия.
Окисление глюкозы сопровождается ее расщеплением, и поэтому такой процесс принято называть гликолизом (от греч. «гликис» - сладкий и «лизис» - расщепление). Анаэробный гликолиз - это неполное окисление глюкозы без участия кислорода, когда акцептором электронов служит НАД+, тогда как полный аэробный гликолиз происходит при участии кислорода.
На первом этапе гликолиза шестиуглеродная молекула глюкозы - С6Н12О6 расщепляется на две трехуглеродные молекулы пировиноградной кислоты (ПВК) - С3Н4О3. При этом происходит лишь частичное окисление глюкозы с потерей четырех атомов водорода (сравните: две молекулы ПВК С3Н4О3 * 2 = С6Н8О6 и глюкозу - С6Н12О6). За счет электронов и протонов водорода восстанавливаются две молекулы НАД+,енесенных с высоких энергетических уровней глюкозы на более низкий уровень молекул НАД+, происходит образование двух молекул АТФ из АДФ и фосфорной кислоты.
Если кислород в клетке отсутствует или его недостаточно, то две молекулы ПВК, образовавшиеся из глюкозы, восстанавливаются за счет двух НАД*Н до молочной кислоты: 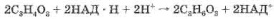
Этим завершается анаэробный гликолиз. В результате бескислородного неполного окисления глюкозы образуются всего только две молекулы АТФ.
Если в клетке уже имеется или же в нее начинает поступать кислород, то ПВК не восстанавливается до молочной кислоты, а переносится в митохондрии, где подвергается при участии кислорода полному окислению до СО2 и Н2О (как при горении). Энергетический выигрыш аэробного гликолиза, выраженный в молекулах АТФ, значительно выше, чем при анаэробном гликолизе.
8888888.. Регуляция гликолиза в целом, сразу для всего организма, происходит под действием гормонов, которые, влияя через молекулы вторичных посредников, изменяют внутриклеточный метаболизм.
Важное значение в стимуляции гликолиза принадлежит инсулину. Глюкагон и адреналин являются наиболее значимыми гормональными ингибиторами гликолиза .
Инсулин стимулирует гликолиз через:
1активацию гексокиназной реакции;
2стимуляцию фосфофруктокиназы;
3стимуляцию пируваткиназы.
Также на гликолиз влияют и другие гормоны. Например, соматотропин ингибирует ферменты гликолиза , а тиреоидные гормоны являются стимуляторами.
Регуляция гликолиза осуществляется через несколько ключевых этапов. Реакции , катализируемые гексокиназой (1), фосфофруктокиназой (3) и пируваткиназой (10) отличаются существенным уменьшением свободной энергии и являются практически необратимыми, что позволяет им быть эффективными точками регуляции гликолиза .
У некоторых видов микроорганизмов, живущих без доступа кислорода, имеет место только анаэробный гликолиз (например, у метанобразующих бактерий, которые живут глубоко в иле болот или озер). Мы можем короткое время обходиться без кислорода, и тогда клетки получают энергию за счет анаэробного гликолиза. Быстрый бег даже на короткие дистанции может вызвать у нас одышку. Это молочная кислота, которая образовалась при недостатке кислорода, возбуждает дыхательный центр и заставляет нас часто дышать, чтобы насытить кровь кислородом, окислить молочную кислоту, восстановить аэробный гликолиз.
Аэробный распад глюкозы
Энергетическое значение аэробного распада глюкозы.
В аэробном гликолизе образуется 10 моль АТФР на 1 моль глюкозы . Так, в реакциях 7, 10 образуется 4 моль АТФ путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТФ (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования.
Баланс аэробного гликолиза.
Суммарный эффект аэробного гликолиза составляет 8 моль АТФ, так как в реакциях 1 и 3 используется 2 моль АТФ. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТФ (по 15 моль на каждую молекулу пирувата. Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТФ.
Анаэробный и аэробный гликолиз энергетически неравноценны. Образование двух моль лактата из глюкозы сопровождается синтезом всего двух моль АТФ, потому что NADH, полученный при окислении глицероальдегидфосфата, не используется дыхательной цепью, а акцептируется пируватом
Анаэробный гликолиз, несмотря на небольшой энергетический эффект, является основным источником энергии для скелетных мышц в начальном периоде интенсивной работы, т. е. в условиях, когда снабжение кислородом ограничено. Кроме того, зрелые эритроциты извлекают энергию за счет анаэробного окисления глюкозы, потому что не имеют митохондрий.
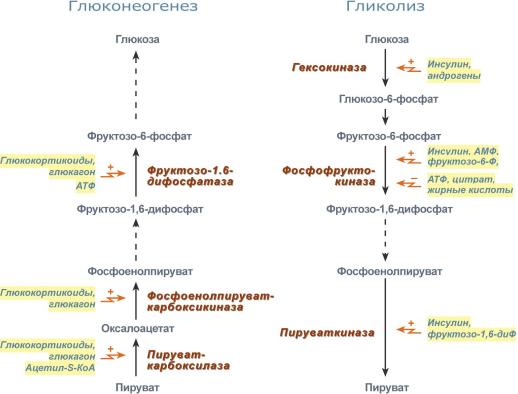
10…… Глюконеогенез— процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободных аминокислот, молочной кислоты, глицерина. Свободные жирные кислоты у млекопитающих для глюконеогенеза не используются.
Стадии глюконеогенеза
1Превращение пирувата в оксалоацетат (фермент пируваткарбоксилаза)
2Превращение оксалоацетата в фосфоенолпируват (фермент фосфоенолпируваткарбоксикиназа)
3Превращение фруктозо-1,6-дифосфата в фруктозо-6-фосфат (фермент фруктозо-1,6-дифосфатаза)
4Превращение глюкозо-6-фосфата в глюкозу (фермент глюкозо-6-фосфатаза)
Суммарное уравнение глюконеогенеза: 2 CH3COCOOH + 4ATP + 2GTP + 2NADH.H+ + 6 H2O = C6H12O6 + 2NAD + 4ADP + 2GDP + 6Pn[1].
Роль в организме
При голодании в организме человека активно используются запасы питательных веществ (гликоген, жирные кислоты). Они расщепляются до аминокислот, кетокислот и других неуглеводных соединений. Большая часть этих соединений не выводится из организма, а подвергаются реутилизации. Вещества транспортируются кровью в печень из других тканей, и используются в глюконеогенезе для синтеза глюкозы — основного источника энергии в организме. Таким образом при истощении запасов организма, глюконеогенез является основным поставщиком энергетических субстратов.
Регуляция гликолиза
В печени кроме пары противоположных процессов синтеза и распада гликогена, свойственной многим клеткам, есть и другая пара противоположных процессов — гликолиз и глюконеогенез (в других органах — только гликолиз ). Как и в случаесинтеза и распада гликогена, регуляция гликолиза и глюконеогенеза связана с ритмом питания, с гормонами инсулином и глюкагоном и с фосфорилированием-де-фосфорилированием ферментов. Переключение печени с гликолиза на глюконеогенез и обратно происходит главным образом в результате изменений активности ферментов, образующих I и II субстратные циклы. Глюконеогенез включается в постабсорбтивном состоянии, когда концентрация глюкагона в крови повышена и, следовательно, активирована аденилатциклазная система и протеинкиназа А.
В регуляции I цикла основная роль принадлежит пируваткиназе, фосфорилиро-ванная форма которой неактивна, а дефосфорилированная — активна (рис. 9.31). Следовательно, гликолитическая реакция ФЕП -* пируват ускоряется при пищеварении и замедляется в постабсорбтивном состоянии. Что касается реакций этого цикла, связанных с глюконеогенезом (пируват -+ оксалоацетат -+ ФЕП), то, по всей вероятности, они с определенной скоростью происходят при любых состояниях. Это может быть связано с необходимостью поддержания в клетке определенной концентрации оксалоацетата, поскольку он участвует во многих важных процессах, включая цитратный цикл.
. 
13…..
Цикл Кори — совокупность биохимических ферментативных процессов транспорта лактата из мышц в печень , и дальнейшего синтеза глюкозы из лактата , катализируемое ферментами глюконеогенеза.
При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах или мышцах ) глюкоза вступает на путь анаэробного гликолиза с образованием лактата . Лактат не может далее окисляться, он накапливается (при его накоплении в мышцах раздражаются чувствительные нервные окончания, что вызывает характерное жжение в мышцах ). С током крови лактат поступает в печень . Печень является основным местом скопления ферментов глюконеогенеза (синтез глюкозы из неуглеводных соеднений), и лактат идет на синтез глюкозы.
Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируват подвергается окислительному декарбоксилированию или может подвергаться брожению.
