Бензойная кислота глицин гиппуровая кислота
Билет№24
1. В 1880 году Н.И. Лунин предположил существование каких-то веществ, без которых не может быть полноценного питания. В 1882 году Такаки , отметил что при добавлении в пищу свежих фруктов и овощей не развивается болезнь бери-бери. В 1896 году Эйкман отметил разницу между заболеваниями бери-бери (у кур) при питании полированном и очищенном рисом. В 1912 году Казимир Функ выделил из оболочки риса вещества, предохраняющие от заболеваемости бери-бери, и назвал его «витамин» (вита - жизнь, содержит - амин).
Витамины – это органические минорные вещества пищи, которые выполняют регуляторную роль в обмене веществ. Витамины изучали: Эйкман, Функ (выделил первый витамин), Н.И. Лунин, К.А. Сосин и другие.
Функции витаминов:
а) ко-ферментная (например, витамин РР – входит в качестве ко-фермента в фермент дегидрогеназу; В6 – ко-фермент фермента аминотрансферазы и т.д.);
б) являются аллостерическими активаторами некоторых ферментов;
в) входят в состав специальных белков (например, витамин А – в состав белка родопсина
г) выполняют антирадикальную роль (например, витамин Е,С) и т.д.
Номенклатура витаминов.
1) Буква латинского алфавита.
2) По химической природе (например, витамин А – ретинол).
3) Добавляется приставка «анти-» к заболеванию, которое развивается при отсутствии этого витамина (например, витамин А называют антиксерофтальмическим – препятствующий сухости глаза, витамин Д –антирахитический и т.д.).
Классификация витаминов.
1) Жирорастворимые (А, Д, Е, К).
2) Водорастворимые (В1, В2, В6, В12, РР, С и т.д.).
3) Витаминоподобные вещества (частично синтезируются в организме и обладают витаминными свойствами) – это холин, липоевая кислота, оротовая кислота, убихинон, линолевая кислота, линоленовая кислота, витамин У (или латинское U), инозит, витамин В12 и т.д.
Обмен витаминов.
1) Всасывание витаминов происходит с помощью специальных белков (В12 требует присутствия внутреннего фактора (гликопротеид) секретируемого желудком).
2) Транспорт витаминов (водорастворимые витамины переносятся плазмой, нерастворимые - белками).
3) Встраивание в состав ко-ферментов.
4) Выделение продуктов обмена.
При нарушении обмена витаминов развиваются различного рода патологические состояния – авитаминоз, гиповитаминоз, гипервитаминоз. Причины развития таких состояний могут быть эндогенные (нарушение всасывания, транспорта и т.д.) или экзогенные (недостаточное поступление витаминов в организм, или полное отсутствие их в потребляемой пище, или , наоборот, чрезмерно большое количество витаминов, поступающие в организм). Экзогенные авитаминозы называют алиментарными, а эндогенные – вторичными. Соответственно экзогенные авитаминозы лечат, назначая витамины (через рот), а эндогенные – парентеральным введением витаминов в организм.
При попадании в толстый кишечник,АК под действием гнилостной микрофлоры, претерпевают различные превращения, образуя достаточно токсичные продукты – фенол, индол, скатол, крезол, сероводород, метилмеркаптан и т.д. и менее токсичные – спирты, амины, кетокислоты и т.д.. Так, в процессе распада серусодержащих АК (цистина, цистеина и метионина) в кишечнике образуется сероводород (Н2S), этилмеркаптан (C2H5SH), метилмеркаптан (CH3SH). Основные АК (орнитин, лизин) подвергаются процессу декарбокалирования с образованием аминов, соответственно путресцина и кадаверина. Из ароматических АК при карбоксилировании могут образоваться соответствующие амины (например, триптамин, тирамин), а при разрушении белковых цепей образуются токсичные: фенол, крезол, скатол, индол.
 OH OH OH
OH OH OH
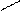

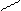
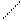



 | | |
| | |












 CH2-CH-COOH
CH2-CH-COOH
 ] ] фенол NH2
] ] фенол NH2
CH2-CH2-COOH CH3 триптофан
 крезол
крезол
NH2
 тирозин
тирозин













 СН3 OH
СН3 OH
 |  | ||||||||||||||
 | |||||||||||||||
 |  | |  |  | |||||||||||
N N N
Скатол индол индоксил
В толстом кишечнике образуется и достаточное количество NH3, который поступает в печень и там различными путями обезвреживается, так как при значительной концентрации обладает токсическим действием. Поэтому больным с поражением печени не рекомендована диета с большим содержанием белков. После всасывания все эти продукты по воротной вене попадают в печень, где подвергаются обезвреживанию путем связывания с серной или глюкуроновой кислотой с образованием так называемых «парных» кислот (например, фенолсерная, индоксилсерная и т. д.), которые выделяются с мочой. Как правило, в реакции вступают активные формы серной и глюкуроновой кислот, соответственно ФАФС (фосфоаденозинфосфосульфат) и УДФГК (урединфосфатглюкуроновая кислота).

 NH2 О
NH2 О



 COOH ||
COOH ||






 N N H O H HN
N N H O H HN

 O OH O
O OH O


 N N HO OH H || O N
N N HO OH H || O N

 HO-S-O-P-OH2C O O-P-OH
HO-S-O-P-OH2C O O-P-OH
 || | H H H H H OH |
|| | H H H H H OH |
O OH O
O OH |

 || HO- P-OH2C O
|| HO- P-OH2C O
HO-P – O ||

 | O H H H
| O H H H
OH H
ФАВС OH OH
УДФГК
Индол (как и скатол) окисляется в индоксил (скатоксил) которые и взаимодействуют либо с ФАФС, либо с УДФГК. Так, индол образует эфир серной кислоты, калиевая соль которой называется животный индикан (выводится с мочой), по его количеству можно судить о скорости гниения белков в кишечнике и о функциональном состоянии печени. О детоксикационной функции печени можно судить по скорости образования и выделения с мочой гиппуровой кислоты, после приема бензойной кислоты.
СООН H2N CO – NH




 | - H2O |
| - H2O |


 + CH2 CH2
+ CH2 CH2
| |
COOH COOH
Бензойная кислота глицин гиппуровая кислота
3.Чрезвычайно большое значение в регуляции углеводного обмена принадлежит гормонам. В ответ на различные отклонения, например, низкий уровень сахара в крови или секрецию гормонов гипоталамуса, в мозговом слое надпочечников синтезируется адреналин. Адреналин с током крови переносится в ткани-мишени, преимущественно в мышцы. Его действие на мышечные клетки приводит к увеличению потребления глюкозы в процессе гликолиза, что улучшает снабжение сокращающихся мышц энергией. Действие его осуществляется с помощью так называемого «регуляторного каскада». На поверхности клетки адреналин вступает в комплекс со специфическим белком R , при этом происходит конформационное изменение аденилатциклазы встроенной в мембрану, в результате которого она активируется и катализирует процесс перехода АТФ в цАМФ. В свою очередь, цАМФ активирует протеинкиназу, ко-торая участвует в АТФ-зависимом фосфорилировании киназы фосфорилазы. В результате этого киназа- фосфорилазы катализирует процесс перехода неактивной формы фосфорилазы «В» в активную форму- фосфорилаза «А». Тем самым усиливается распад гликогена, увеличивается содержание глюкозы, которая при распаде может дать большое количество АТФ . Помимо указанного пути активирования аденилатциклазы, повысить активность указанного результата можно и другим путем. Ионы Са++ способны связываться с белком мембраны кальмодулином. Образовавшийся комплекс вызывает конформационные изменения аденилатциклазы, повышая активность этого фермента. Как известно, при мышечном сокращении происходит значительный выброс ионов Са++. Таким образом, само мышечное сокращение стимулирует процессы гликолиза.
 Гормон пептидной природы - глюкагон. Генерация глюкагона происходит под действием адреналина. a-клетками островков Лангерганса поджелудочной железы в ответ на низкое содержание глюкозы в крови. Мишенью для глюкогона являются, в первую очередь, клетки печени. Под действием глюкогена печень вырабатывает большое количество глюкозы, которая затем с кровью переносится в другие ткани. Такой эффект достигается путем повышения образования глюкозо-6-фосфата в ходе цАМФ –зависимого регуляторного каскада, в некоторой степени сходного с адреналиновым каскадом. Глюкогоновый каскад запускается при связывании глюкогона со специфическим мембранным рецепторным белком, это приводит к активации аденилатциклазы и повышает уровень цАМФ, что активирует гликоген-фосфорилазу печени и ингибирует гликоген-синтазу. В результате повышается скорость образования Г-1-Ф и , следовательно, Г-6-Ф. Таким образом, в этом отношении глюкогон по своему действию идентичен адреналину-они оба активируют гликолиз посредством цАМФ. Однако, недавно было показано, что в печени глюкогоновый каскад включает также другой регуляторный компонент-Фруктозу-2,6-дифосфат. Его открыли в 1980г., т.к. он присутствует в очень малых количествах. Фактически, при определенных условиях его клеточная концетрация близка к О.
Гормон пептидной природы - глюкагон. Генерация глюкагона происходит под действием адреналина. a-клетками островков Лангерганса поджелудочной железы в ответ на низкое содержание глюкозы в крови. Мишенью для глюкогона являются, в первую очередь, клетки печени. Под действием глюкогена печень вырабатывает большое количество глюкозы, которая затем с кровью переносится в другие ткани. Такой эффект достигается путем повышения образования глюкозо-6-фосфата в ходе цАМФ –зависимого регуляторного каскада, в некоторой степени сходного с адреналиновым каскадом. Глюкогоновый каскад запускается при связывании глюкогона со специфическим мембранным рецепторным белком, это приводит к активации аденилатциклазы и повышает уровень цАМФ, что активирует гликоген-фосфорилазу печени и ингибирует гликоген-синтазу. В результате повышается скорость образования Г-1-Ф и , следовательно, Г-6-Ф. Таким образом, в этом отношении глюкогон по своему действию идентичен адреналину-они оба активируют гликолиз посредством цАМФ. Однако, недавно было показано, что в печени глюкогоновый каскад включает также другой регуляторный компонент-Фруктозу-2,6-дифосфат. Его открыли в 1980г., т.к. он присутствует в очень малых количествах. Фактически, при определенных условиях его клеточная концетрация близка к О.
Инсулин синтезируется b-клетками островков Лангерганса поджелудочной железы в ответ на повышение содержания глюкозы в крови. Клетками –мишенями для инсулина в первую очередь являются клетки мышц, печени и жировой ткани. В клетках каждого типа инсулин вначале связывается со специфическими белковыми рецепторами клеточной мембраны. В результате связывания стимулируется транспорт глюкозы внутрь клеток,а затем стимулируется утилизация внутриклеточной глюкозы в процессе синтеза гликогена, а также в процессе белкового и липидного синтеза. В результате происходит понижение содержания глюкозы в крови до нормального уровня, –а это сигнал для поджелудочной железы остановить синтез инсулина. Действие инсулина до конца еще не ясно. Возможно, стимуляция синтеза гликогена вызвана активацией цАМФ-фосфодиэстеразы инсулином. Это фермент, который разрушает цАМФ. При этом снижается содержание в клетке цАМФ, активируется гликоген-синтаза, что стимулирует синтез гликогена. Оказывают влияние на обмен углеводов и другие гормоны. Так, соматотропин (гормон роста), синтезируемый передней долей гипофиза, тормозит сек-рецию инсулина. При гиперсекреции соматотропина возникает так называемый гипофизарный диабет. Соматостатин, вырабатываемый Д- клетками поджелудочной железы, ингибирует секрецию соматотропина и сложным образом влияет на секрецию инсулина и глюкогона. Он используется при лечении некоторых форм сахарного диабета. Влияют на обмен углеводов и гормоны коры надпочечников. Кортизол (важнейший представитель глюкокортикондов) стимулирует процесс глюко-неогенеза из аминокислот и способствует накоплению гликогена в печени. Он также повышает уровень глюкозы в крови и снижает использование глюкозы в периферических тканях. Определенное влияние оказывают и гормоны щитовидной железы. Тироксин, связывается со специфическими рецепторными белками, которые обеспечивают проникновение тироксина в клеточное ядро, где он, связываясь со специфическими генами, резко повышает синтез определенных ферментов и ферментных систем.