Активный транспорт и его виды.
Транспорт веществ через биологические мембраны (продолжение).
Активный транспорт и его виды.
Активный транспорт – это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением (рис. 5.10).

Рис. 5.10. Схема активного транспорта.
Активный транспорт в мембране сопровождается ростом энергии Гиббса, он не может идти самопроизвольно, а только в сопряжении с процессом гидролиза АТФ, то есть за счет затраты энергии. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, давления и другие процессы, поддерживающие жизненные функции. Активный транспорт удерживает организм в неравновесном состоянии.
Существование активного транспорта впервые было доказано в опытах Уссинга (1949 г.) на примере переноса ионов натрия через кожу лягушки (рис. 5.11).
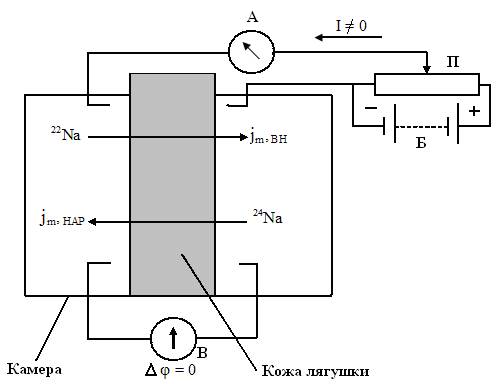
Рис. 5.11. Схема опыта Уссинга (А – амперметр, В – вольтметр, Б – батарея, П – потенциометр).
Экспериментальная камера Уссинга, заполненная раствором Рингера, разделена на две части свежеизолированной кожей лягушки. На рис. 6.11 слева – наружная мукозная поверхность кожи, справа – внутренняя серозная. Уссинг наблюдал потоки ионов натрия через кожу лягушки: слева направо от наружной к внутренней поверхности и справа налево от внутренней к наружной поверхности.
Из уравнения Теорелла, которое описывает пассивный транспорт, следует уравнение Уссинга – Теорелла для отношения потоков в случае пассивного транспорта:
jm, ВН/jm, НАР = (СНАР/СВН) ех,
где: х = (Z F )/R Т.
На коже лягушки, разделяющей раствор Рингера, возникает разность потенциалов = ВН – НАР. Внутренняя сторона кожи имеет положительный потенциал по отношению к наружной. В установке Уссинга имеется блок компенсации напряжения, с помощью которого устанавливается разность потенциалов на коже лягушки, равная нулю, что контролируется вольтметром. Кроме этого, поддерживается одинаковая концентрация ионов с наружной и внутренней стороны СНАР = СВН.
При этих условиях, если бы перенос натрия через кожу лягушки определялся только пассивным транспортом, то согласно уравнению Уссинга – Теорелла jm, ВН и jm, НАР были бы равны друг другу: jm, ВН = jm, НАР. Суммарный поток через мембрану был бы равен нулю.
Однако, обнаружено с помощью вольтметра, что в условиях опыта (при отсутствии градиентов электрического потенциала и концентрации) через кожу лягушки течет электрический ток I, следовательно, происходит односторонний перенос заряженных частиц. Ток протекает через кожу от внешней стороны к внутренней.
С помощью меченых атомов было установлено, что поток натрия внутрь больше потока его наружу: jm, ВН > jm, НАР. Для этого в левый раствор экспериментальной камеры были включены радиоактивные изотопы 22Na, а в правый – 24Na. Изотоп 22Na распадается с излучением жестких – квантов. Распад 24Na сопровождается мягким – излучением. Регистрация – и – излучения показала, что поток 22Na больше, чем поток 24Na.
Этот эксперимент свидетельствует, что перенос ионов натрия через кожу лягушки не подчиняется уравнению пассивного транспорта, значит, имеет место активный перенос.
Перенос иона в процессе активного транспортаосуществляется транспортными АТФ–азами, энергозависимой лимитирующей стадией которого является гидролиз АТФ. Так происходит первично-активныйтранспорт.
Если же перенос веществ обеспечивает энергия, обусловленная градиентом ионов, созданным в ходе первично–активноготранспорта, то это вторично–активныйтранспорт. С его помощью осуществляется перенос сахаров и аминокислот за счет энергии, обусловленной градиентом ионов, чаще натрия, созданного первично-активным транспортом.
Функциональное значение активного транспорта определяется тем, что он осуществляет неравномерное распределение ионов между клеткой и средой – для большинства тканей концентрация внутриклеточного калия больше, чем внеклеточного, а концентрация внутриклеточного натрия меньше, чем внеклеточного. Поддержание постоянного ионного состава клетки обеспечивает ионный гомеостаз, необходимый для осуществления жизненно важных градиентзатратных процессов.
Транспортные АТФ–азы прокариотических и эукаритических клеток делятся на 3 типа: P–тип, V–тип, F–тип.
АТФ–азы P–типаявляются примером осуществления обязательной стадии фосфорилирования и дефосфорилирования, сопряженной со структурными переходами белков, то есть образования ковалентного фосфорилированного промежуточного продукта (фосфомедиата).
К АТФ–азам цитоплазматической мембраны этого типа относятся:
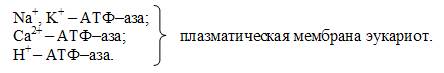
Внутриклеточные АТФ-азы P–типа:
Ca2+ – АТФ–аза эндо–(сарко) плазматического ретикулума эукариот.
К+ – АТФ-аза наружных мембран прокариот. Устроены довольно просто, действуют как насос.
АТФ–азы V–типанаходятся в мембранах в вакуолях дрожжей, в лизосомах, эндосомах, секреторных гранулах животных клеток (Н+ – АТФ–азы).
АТФ–азы F–типанаходятся в мембранах бактерий, в хлоропластах, митохондриях. Они образованы:
1.Водорастворимой частью – F1, которая состоит из нескольких субъединиц и обладает каталитической активностью.
2.Гидрофобная часть Fо, участвующей в транслокации водорода. Особенностью АТФ–азы F–типаявляется способность синтезировать АТФ за счет переноса протонов по градиенту концентрации.