Эрнст Генрих Филипп Август Геккель
Эрнст Генрих Филипп Август Геккель (нем. Ernst Heinrich Philipp August Haeckel; 1834—1919) — немецкий естествоиспытатель и философ. Автор терминов питекантроп и экология.
В молодости, начиная с 1852 года, изучал медицину и естествознание в Берлинском, Вюрцбургском и Венском университетах. В 1858 году сдал экзамен и получил диплом врача, хотя медицинской практикой позже он никогда не занимался. В это время более всего его интересуют исследования живой природы, в первую очередь зоология и сравнительная микроскопическая анатомия.[1] В 1859 году Геккель участвовал в научной экспедиции в Италию, во время которой во Флоренции он приобрёл мощный микроскоп в мастерской известного натуралиста и оптика Амичи.
Вскоре Геккель занялся изучением морского планктона в Мессинском проливе. Исследования велись в течение шести месяцев с помощью нового микроскопа. В результате было обнаружено 120 новых видов радиолярий[1]. Данное направление исследований стало для него одним из основных до конца жизни. Во времена Геккеля было известно несколько сотен видов радиолярий, в современной науке известно более 5000. Эрнст Геккель представил доклад по теме радиолярий в 1860 году, на тридцать пятом съезде Общества немецких естествоиспытателей и врачей. В 1862 году, в возрасте двадцати восьми лет, Геккель был назначен адъюнкт-профессором, позже приват-доцентом в университете Йены.
Сильнейшее воздействие на Геккеля оказали дарвиновские идеи. В 1863 он выступил с публичной речью о дарвинизме на заседании Немецкого научного общества, а в 1866 вышла его книга «Общая морфология организмов» («Generelle Morphologie der Organismen»). Спустя два года появилась «Естественная история миротворения» («Natürliche Schöpfungsgeschichte»; рус. перевод 1914), где развиваемый им эволюционный подход излагался в более популярной форме, а в 1874 Геккель опубликовал работу «Антропогения», или «История развития человека» («Anthropogenie», или «Entwickelungsgeschichte des Menschen»; русский перевод 1919 года), в которой обсуждались проблемы эволюции человека. Ему принадлежит мысль о существовании в историческом прошлом формы, промежуточной между обезьяной и человеком, что до сих пор не подтверждено.
Геккель разработал теорию происхождения многоклеточных (так называемая теория гастреи) (1866), сформулировал биогенетический закон, согласно которому в индивидуальном развитии организма как бы воспроизводятся основные этапы его эволюции, построил первое генеалогическое древо животного царства.
Теория гастреи принесла ему известность и признавалась большинством учёных до сравнительно недавнего времени. В настоящее время эволюционисты, наряду с теорией гаструлы, рассматривают как хорошо обоснованную теорию фагоцителлы, предложенную И. И. Мечниковым в 1879-86 гг., а также теорию синзооспоры и первичной седентарности многоклеточных, разработанную А. А. Захваткиным, и другие.
Продолжая свои зоологические исследования в лаборатории и в ходе экспедиций на остров Мадейра, на Цейлон, в Египет и Алжир, Геккель публикует монографии по радиоляриям, глубоководным медузам, сифонофорам, глубоководным рыбам-удильщикам, а также свой последний систематический труд — внушительную «Систематическую филогению» («Systematische Philogenie», 1894—1896; русский перевод 1899 года).
Для зоологических исследований предпринимал поездки на Гельголанд и в Ниццу, работал в Неаполе и Мессине. Путешествовал в Лиссабон, на Мадейру, Тенерифе, Гибралтар, в Норвегию, Сирию и Египет, на Корсику, Сардинию и Цейлон. Геккель был одним из первых германских зоологов, поддержавших теорию Дарвина. Опираясь на эту теорию и на данные эмбриологии, Геккель сделал попытку дать рациональную систему животного царства, основанную на филогенезе животных. Особенное внимание обратил Геккель на важное значение истории развития индивидуального, или онтогения, для вопроса о происхождении самого вида или его филогении. Исходной точкой для взглядов Геккеля послужила стадия эмбрионального развития, названная Геккелем гаструлой. Геккель полагал, что эта стадия повторяет собой общего прародителя всех животных. Этого предполагаемого прародителя Геккель назвал гастреей и старался объяснить, каким образом из неё развились различные типы животного царства. Учение о гастрее позднее было признано ошибочным.
Геккель представил генеалогию растительного царства, начиная от самых простейших форм, протистов, до сростнолепестных, считаемых за самые развитые и совершенные формы.
Реакция фотосинтеза:
образование живыми растительными клетками органических веществ, таких, как сахара и крахмал, из неорганических - из СО2 и воды - с помощью энергии света, поглощаемого пигментами растений. Это процесс производства пищи, от которого зависят все живые существа - растения, животные и человек. У всех наземных растений и у большей части водных в ходе фотосинтеза выделяется кислород. Некоторым организмам, однако, свойственны другие виды фотосинтеза, проходящие без выделения кислорода. Главную реакцию фотосинтеза, идущего с выделением кислорода, можно записать в следующем виде:

К органическим веществам относятся все соединения углерода за исключением его оксидов и нитридов. В наибольшем количестве образуются при фотосинтезе такие органические вещества, как углеводы (в первую очередь сахара и крахмал), аминокислоты (из которых строятся белки) и, наконец, жирные кислоты (которые в сочетании с глицерофосфатом служат материалом для синтеза жиров). Из неорганических веществ для синтеза всех этих соединений требуются вода (Н2О) и диоксид углерода (СО2).
Роль фотосинтеза. Суммарный итог химических реакций фотосинтеза может быть описан для каждого из его продуктов отдельным химическим уравнением. Для простого сахара глюкозы уравнение имеет следующий вид:

Уравнение показывает, что в зеленом растении за счет энергии света из шести молекул воды и шести молекул диоксида углерода образуется одна молекула глюкозы и шесть молекул кислорода. Глюкоза - это лишь один из многих углеводов, синтезируемых в растениях. Ниже приведено общее уравнение для образования углевода с n атомами углерода в молекуле:

Уравнения, описывающие образование других органических соединений, имеют не столь простой вид. Для синтеза аминокислоты требуются дополнительные неорганические соединения, как например при образовании цистеина:
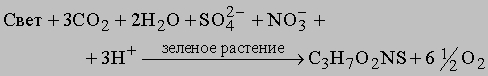
Роль света как реагента в процессе фотосинтеза легче доказать, если обратиться к другой химической реакции, а именно к горению. Глюкоза - одна из субъединиц целлюлозы, главного компонента древесины. Горение глюкозы описывается следующим уравнением:

Это уравнение представляет собой обращение уравнения фотосинтеза глюкозы, если не считать того, что вместо световой энергии выделяется главным образом тепло. По закону сохранения энергии, если при горении энергия выделяется, то при обратной реакции, т.е. при фотосинтезе, она должна поглощаться.
Круговорот углерода
Углерод в биосфере часто представлен наиболее подвижной формой - углекислым газом. Источником первичной углекислоты биосферы является вулканическая деятельность, связанная с вековой дегазацией мантии и нижних горизонтов земной коры. Миграция углекислого газа в биосфере Земли протекает двумя путями. Первый путь заключается в поглощении его в процессе фотосинтеза с образованием органических веществ и в последующем захоронении их в литосфере в виде торфа, угля, горных сланцев, рассеянной органики, осадочных горных пород. Так, в далекие геологические эпохи сотни миллионов лет назад значительная часть фотосинтезируемого органического вещества не использовалась ни консументами, ни редуцентами, а накапливалась и постепенно погребалась под различными минеральными осадками. Находясь в породах миллионы лет, этот детрит под действием высоких температур и давления (процесс метаморфизации) превращался в нефть, природный газ и уголь, во что именно - зависело от исходного материала, продолжительности и условий пребывания в породах. Теперь мы в огромных количествах добываем это ископаемое топливо для обеспечения потребностей в энергии, а сжигая его, в определенном смысле завершаем круговорот углерода. Если бы ни этот процесс в истории планеты, вероятно, человечество имело бы сейчас совсем другие источники энергии, а может быть и совсем другое направление развития цивилизации.

По второму пути миграция углерода осуществляется созданием карбонатной системы в различных водоемах, где CO2 переходит в H2CO3, HCO31-, CO32-. Затем с помощью растворенного в воде кальция (реже магния) происходит осаждение карбонатов CaCO3 биогенным и абиогенным путями. Возникают мощные толщи известняков. Наряду с этим большим круговоротом углерода существует еще ряд малых его круговоротов на поверхности суши и в океане. В пределах суши, где имеется растительность, углекислый газ атмосферы поглощается в процессе фотосинтеза в дневное время. В ночное время часть его выделяется растениями во внешнюю среду. С гибелью растений и животных на поверхности происходит окисление органических веществ с образованием CO2. Особое место в современном круговороте веществ занимает массовое сжигание органических веществ и постепенное возрастание содержания углекислого газа в атмосфере, связанное с ростом промышленного производства и транспорта.
Круговорот кислорода

Кислород - наиболее активный газ. В пределах биосферы происходит быстрый обмен кислорода среды с живыми организмами или их остатками после гибели. В составе земной атмосферы кислород занимает второе место после азота. Господствующей формой нахождения кислорода в атмосфере является молекула О2. Круговорот кислорода в биосфере весьма сложен, поскольку он вступает во множество химических соединений минерального и органического миров. Свободный кислород современной земной атмосферы является побочным продуктом процесса фотосинтеза зеленых растений и его общее количество отражает баланс между продуцированием кислорода и процессами окисления и гниения различных веществ. В истории биосферы Земли наступило такое время, когда количество свободного кислорода достигло определенного уровня и оказалось сбалансированным таким образом, что количество выделяемого кислорода стало равным количеству поглощаемого кислорода.
Круговорот серы
Сера является важным составным элементом живого вещества. Большая часть ее в живых организмах находится в виде органических соединений. Кроме того, сера входит в состав некоторых биологически активных веществ: витаминов, а также ряда веществ, выступающих в качестве катализаторов окислительно-восстановительных процессов в организме и активизирующих некоторые ферменты. Сера представляет собой исключительно активный химический элемент биосферы и мигрирует в разных валентных состояниях в зависимости от окислительно-восстановительных условий среды. Среднее содержание серы в земной коре оценивается в 0,047 %. В природе этот элемент образует свыше 420 минералов.
В изверженных породах сера находится преимущественно в виде сульфидных минералов: пирита , пирронита , халькопирита , в осадочных породах содержится в глинах в виде гипсов, в ископаемых углях - в виде примесей серного колчедана и реже в виде сульфатов. Сера в почве находится преимущественно в форме сульфатов; в нефти встречаются ее органические соединения. В связи с окислением сульфидных минералов в процессе выветривания сера в виде сульфатиона переносится природными водами в Мировой океан. Сера поглощается морскими организмами, которые богаче ее неорганическими соединениями, чем пресноводные и наземные.
БИОМЫ
Тундра
Северную полярную область опоясывает тундра — холодная безлесная равнина, занимающая огромные пространства в Евразии, Северной Америке и Гренландии; Марко Поло охарактеризовал ее как «область, недоступную из-за болот и льда». В Южном полушарии аналогичной зоны нет, так как оно занято в основном океаном. В тундре лимитирующими факторами среды являются низкие температуры, малое количество осадков и короткий вегетационный период (около 60 дней). Верхний слой грунта толщиной от нескольких сантиметров до нескольких метров периодически промерзает и оттаивает, а под ним лежит слой многолетней мерзлоты. В связи с этим в низинах дренаж почвы недостаточен и здесь много сырых мест, болот и мелких водоемов. Низменные участки покрыты рыхлым ковром из приземистой растительности и медленно разлагающихся растительных остатков: низкие температуры замедляют деятельность бактерий. Возвышенные участки обычно голые и каменистые. Под действием морозов происходит постоянное перемещение обломков грунта, в результате чего возникают каменные гирлянды; верхний слой почвы растрескивается, образуя полигональные структуры, а оттаявшая почва, сползая по слою мерзлоты, образует валы и солифлюкционные террасы. В этих условиях растениям трудно закрепляться в грунте и выдерживать напор ветра, поэтому даже небольшие различия в топографии земной поверхности (например, различие между подветренной и наветренной сторонами каменных многоугольников) и снежного покрова становятся важными экологическими факторами. Число видов растений невелико, и это обычно приземистые формы. К характерным растениям тундры относятся пушица, наземные виды осок и ситников, мхи, лишайники, карликовые формы вереска, карликовые березы и ивы. Животные приспособлены к холоду, длинной зиме, короткому лету и скудному растительному покрову; почти все виды имеют циркумполярное распространение, что связано с наличием почти непрерывной полосы суши вокруг полюса, хотя в Северной Америке некоторым евразийским видам соответствуют иные, но близкие к ним виды. Например, в Старом Свете встречаются северный олень и росомаха, а в Новом Свете — карибу (дикий северный олень) и американская росомаха. Из других животных можно упомянуть мускусного овцебыка, полярного зайца, песца, длиннохвостого аляскинского суслика, лемминга, тундряную куропатку и конька, а также многочисленных песочников и ржанок (в сырых местах). Беспозвоночных в тундре мало, однако летом здесь летают тучи комаров и мошек.
Из-за короткого лета цикл жизни у некоторых беспозвоночных сильно растянут и слагается из нескольких периодов летней активности, разделенных периодами зимней спячки; завершение всего цикла развития иногда занимает несколько лет. Напротив, у позвоночных наблюдается тенденция к ускоренному развитию. Например, северная форма дрозда Тurdus migratorius кормит своих птенцов в течение 21 ч в сутки, так что период роста занимает у них всего 8, а не 13 дней, как у южных форм.
Лиственные леса
К югу от хвойных лесов лежит область листопадных лесов умеренной зоны, которые прежде занимали восточную часть Северной Америки, всю Европу, часть Китая, Японии и Австралии и южную часть Южной Америки. В Европе, Северной Америке и на Дальнем Востоке цивилизация достигла наибольшего развития именно в этих районах, и в результате от некогда огромных лесных массивов почти ничего не осталось. Когда в Америку прибыли английские поселенцы, материк был чуть ли не сплошь покрыт лесом; говорили, что «белка могла бы добраться от Атлантического океана до Миссисипи, ни разу не коснувшись земли». А глядя на нынешнюю Европу, никак нельзя представить себе, какие обширные и густые леса покрывали ее в средние века и раньше.
Биом лиственного леса богат и сложен. Он подразделяется на много региональных разновидностей, а число различных древесных пород здесь очень велико. Например, в Северной Америке в переходной зоне между хвойным лесом и восточными листопадными лесами доминируют банксова и норвежская сосны, канадская тсуга, кипарисовик Лавсона, туя, бук, сахарный клен, американская липа, желтая береза, поздняя черемуха и серебристый дуб. Одним из доминирующих видов раньше была величественная веймутова сосна, особенно в области, окружающей Великие озера, где она была уничтожена в результате варварского ведения лесозаготовок в конце XIX века. К важным разновидностям лиственного леса относятся: буково-кленовый лес (на севере центральной части США), кленово-липовый лес (в Висконсине и Миннесоте), лес из дуба с гикори (в западных и южных штатах), дубово-каштановый лес (в Аппалачских горах до массового поражения каштана раком), смешанные мезофитные леса Аппалачского плато (с очень большим числом видов, среди которых доминирует тюльпанное дерево) и сосновые леса на юге. Последние не являются конечной стадией сукцессии; они сохраняются благодаря рубкам и лесным пожарам — без вмешательства этих факторов сосна уступила бы место дубу, гикори и магнолии. Виргинский дуб и магнолия доминируют во Флориде и в других штатах, расположенных у Мексиканского залива.
Восточный листопадный лес занимает область, характеризующуюся мощным слоем плодородной почвы, равномерным распределением осадков и отсутствием резких колебаний температуры. Сезонные изменения хорошо выражены. В этих лесах с пышно развитым травянистым и кустарниковым подлеском обитает очень много птиц и млекопитающих, поэтому здесь трудно выделить какие-то особо характерные виды.
Пустыни
Одну седьмую часть всей поверхности суши занимают области, где дожди не восполняют потерю влаги в результате испарения. Для всех этих областей, называемых пустынями, характерен недостаток воды, хотя они могут отличаться друг от друга во многих других отношениях. Только в северном Чили и в Центральной Сахаре имеются районы, где совсем не бывает дождей. В большинстве других пустынь выпадает некоторое количество осадков, обычно менее 25 см в год. Наиболее обширные пустыни образуют на земном шаре два прерывистых пояса, лежащих приблизительно в зонах северного и южного тропиков. Самая обширная из них, Сахара, занимает около 9 000 000 км2, но эта область неоднородна по ландшафту: лишь около 10% ее занимают песчаные дюны, а в западной части есть даже горы, покрытые снегом. На востоке к Сахаре примыкает Аравийская пустыня, третью часть которой покрывают пески; она отличается полным отсутствием рек, которые начинались бы в ней или пересекали ее. Дальше на восток расположены пустыни Средней Азии, Ирана, Индии, пустыни Такла-Макан и Гоби. В Северной Америке пустыни имеются на западе США и в прилегающей части Мексики (Большой Бассейн, Мохаве, Сонора и Чихуахуа). К югу от экватора расположены Австралийская пустыня, занимающая 44% площади материка, пустыни Патагонии в Аргентине, пустыни Атакама в Чили и Калахари в Африке.
Большинство пустынь отличается жарким климатом, температура воздуха летом доходит до 50° С, а температура почвы может быть примерно на 15 — 25° С выше. Но в некоторых пустынях (например, Гоби) климат холодный из-за большой высоты над уровнем моря. Как правило, наблюдается большой контраст между дневными и ночными и между летними и зимними температурами. В пустыне может быть и очень холодно, и очень жарко. Организмы, обитающие в пустынях, должны приспосабливаться к крайним температурам, к недостатку воды, к сильным ветрам, к выпадающим изредка ливням, а в некоторых областях и к засоленности почвы.
Растения пустынь либо избегают воздействия засухи благодаря своему жизненному циклу, либо обладают засухоустойчивостью. К первым относятся однолетники, которые переживают сухой период в виде семян, а после выпадения достаточного количества осадков прорастают, быстро зацветают, производят семена и затем отмирают. Ко второй группе относятся многолетние растения, которые либо имеют корни, доходящие до подземных вод, либо сбрасывают листву в период засухи, либо запасают в своих тканях огромные количества воды. Многие из таких видов растут около пересыхающих рек или в местах, где имеются подземные запасы воды. У мескитового дерева корни уходят в землю на глубину до 30 м. Растущий в американских пустынях паловерде (Cercidium macrum) испаряет мало воды благодаря очень мелким листьям. Суккулентами называются растения, сохраняющие запасы воды в толстых, мясистых листьях или в боченкообразных стеблях. Взрослое растение кактуса сагуаро может весить до 10 тонн, причем четыре пятых этого веса приходится на воду. Многие растения пустынь покрыты колючками, спасающими их от некоторых растительноядных животных. Лимитирующий фактор в пустыне уже не свет, как в лесу, а вода. Для пустыни характерны такие растения, как кассии, акации, юкки (в том числе древовидная юкка), агавы, кактусы, полынь, креозотовый кустарник, различные молочаи и эфиромасличные кустарники.
Для животных проблему составляют нехватка воды и крайне высокие температуры. Многие обитатели пустынь ведут ночной или подземный образ жизни. Некоторые виды используют метаболическую воду. В пустыне обитает много видов животных; наиболее типичны мешотчатый прыгун, песчанки, тушканчики, сцинки и другие ящерицы, а из хищников — собака динго в Австралии, африканская дикая собака, койоты и карликовые лисицы. В пустынях живет также невероятное множество насекомых.
Влажный тропический лес, Тропический дождевой лес (англ. Tropical rain forest; Moist tropical forest) — биом, лес в экваториальных (влажный экваториальный лес), субэкваториальных и влажнотропических районах с влажным климатом (2000—7000 мм осадков в год). В дополнение к чрезмерному выпадению осадков, влажные тропические леса характеризуются большим количеством постоянных (в противоположность мигрирующим) видов животных и огромным биоразнообразием флоры и фауны.
Во влажных тропических лесах живёт две трети всех видов животных и растений планеты. Предполагается, что миллионы видов животных и растений до сих пор не описаны. Эти леса иногда называют «драгоценностями Земли» и «самой большой аптекой мира», поскольку большое количество природных медицинских средств было найдено здесь. Их ещё также называют «лёгкими Земли», однако это утверждение спорно, поскольку не имеет научного обоснования, так как эти леса либо совсем не вырабатывают кислород, либо вырабатывают его крайне мало[1]. Но следует иметь в виду, что влажный климат способствует эффективной фильтрации воздуха, благодаря конденсации влаги на микрочастицах загрязнений, что оказывает в целом благоприятное воздействие на атмосферу.
Образование подлеска в тропических лесах сильно ограничено во многих местах ввиду недостатка солнечного света на нижнем ярусе. Это позволяет человеку и животным продвигаться по лесу. Если по какой-либо причине лиственный навес отсутствует или ослаблен, нижний ярус быстро покрывается плотной зарослью винограда, кустарников и небольших деревьев — такое образование называется джунглями.
Тайга — биом, характеризующийся преобладанием хвойных лесов (бореальных видов ели, пихты, лиственницы, сосны, в том числе кедровой).
Флора
Тайге свойственно отсутствие или слабое развитие подлеска (так как в лесу мало света), а также однообразие травяно-кустарникового яруса и мохового покрова (зелёные мхи). Виды кустарников (можжевельник, жимолость, смородина и др.), кустарничков (черника, брусника и др.) и трав (кислица, грушанка) немногочисленны как в Евразии, так и в Северной Америке.
На севере Европы (Финляндия, Швеция, Норвегия, Россия) преобладают еловые леса, в Северной Америке (Канада) — еловые леса с примесью лиственницы канадской. Для тайги Урала характерны светлохвойные леса из сосны обыкновенной. В Сибири и на Дальнем Востоке господствует редкостойная лиственничная тайга с подлеском из кедрового стланика, рододендрона даурского и другое.
Фауна
Животный мир тайги богаче и разнообразнее, чем животный мир тундры. Многочисленны и широко распространены: рысь, росомаха, бурундук, соболь, белка и др. Из копытных встречаются северный и благородный олень, лось, косуля; многочисленны зайцы, бурозубки, грызуны: мыши, полёвки, белки обыкновенная и летяга, бурундуки. Из птиц обычны: глухарь, обыкновенный рябчик, кедровка, клесты и др. Для тайги Северной Америки типичны американские виды тех же родов, что и в Евразии.
В таёжном лесу по сравнению с лесотундрой благоприятнее условия для жизни животных. Здесь больше оседлых животных. Нигде в мире, кроме тайги, не водится столько пушных зверей.
В зимний период подавляющее число видов беспозвоночных, все земноводные и пресмыкающиеся, а также некоторые виды млекопитающих погружаются в анабиоз и зимнюю спячку, снижается активность ряда других животных.
Степи умеренной зоны расположены там, где выпадает промежуточное между пустынями и лесами количество осадков (250— 750 мм); существование степей в этих условиях зависит от температуры, сезонного распределения осадков и влагоемкости почвы. Упоминавшиеся уже исследования в рамках МБП показали, что ключевым фактором здесь является влажность почвы, в частности потому, что она лимитирует микробное разложение и возврат в круговорот биогенных элементов. Обширные степные пространства занимают внутренние части Евразийского и Североамериканского континентов, юг Южной Америки (аргентинские пампасы) и Австралию.
В Северной Америке биом степей подразделяется (с востока на запад) на следующие зоны: высокотравные, смешанные, низкотравные и кустисто-злаковые прерии. Эта зональность определяется градиентом количества осадков, который одновременно является и градиентом уменьшения первичной продуктивности. Ниже приводится перечень некоторых наиболее важных видов многолетников, сгруппированных по высоте надземных частей.
1. Высокие злаки (от 150 до 240 см) — бородач (Andropogon gerardi), просо (Panieum virgatum), индейская трава (Sorghastrum nutans) и в низинах Spartina pectinata.
2. Средние злаки (от 60 до 120 см) — малый бородач (Andropogon scoparius), ковыль (Stipa spartea), спороболус (Sporobolus heterolepis), пырей (Agropyron smithii), тонконог (Koeleria cristata), индейский рис (Oryzopsis) и многие другие.
3. Низкие злаки (от 15 до 45 см) — бизонова трава (ВисШое dactyloides), голубая грамова трава (Bouteloua gracilis), другие виды грамовой травы (Bouteloua sp.), интродуцированные мятлик (Роа) и костер (Bromus sp.).
Корни большинства видов глубоко (до 2 м) проникают в почву, и масса корней климаксных многолетников в несколько раз превышает массу их надземных частей. Большое значение имеет форма роста корней. Некоторые из упомянутых видов, например бородач, бизонова трава и пырей, образуют подземные корневища и формируют таким образом дернину. Другие виды, такие, как малый бородач, тонконог, ковыль, кустятся и растут куртинами. Эти две жизненные формы можно найти во всех зонах, но кустовые злаки преобладают в более засушливых областях, где степь переходит в пустыню.
Средиземн. Типа
В ряде работ по флоре и растительности (А. Гри зебах, 1872; М. Амберже. 1930; М. Рикли, 1943—1948: Е. Шмид, 1949) к Средиземноморью относятся не только районы с действительно средиземноморской природой, но и другие районы распространения средиземноморской флоры —центральные плоскогорья Малой Азии, Месопотамия и Иранское нагорье, природа которых существенно отличается от природы берегов Средиземного моря. Пределы распространения средиземноморской растительности различными авторами проводятся по-разному. Только по вопросу о границе этой растительности в Европе существует целая литература (Д. Гурлитт, 1950; Ф. Маркграф, 1952; К. Ре-гель, 1940, 1963, и др.). Но ботанико-географическая средиземноморская область ограничивается более тесными рамками. В. П. Малеев (1948) считает, что она «простирается по островам и побережьям Средиземного моря и сообщающихся с ним морей от Пиренейского полуострова и Марокко на западе до побережий Малой Азии, Сирии и Палестины на востоке. К ней относятся Пиренейский, Апеннинский и большая часть Балканского полуострова; в СССР к этой области принадлежит Горный Крым и Западное Закавказье от Анапы на севере и до границы СССР на юге».
Таким образом, Средиземноморье рассматривается в узком и широком смысле. В первом случае ограничивается регион распространения «классических» средиземноморских ландшафтов, которые занимают территории, примыкающие к Средиземному морю в Евразии и Африке. Этот регион примерно совпадает с ареалами распространения жестколистной растительности и культуры маслины, его границы близки к границам средиземноморской ландшафтной зоны . Именно в его пределах распространены характерные для Средиземноморья природные условия. Поэтому в дальнейшем при упоминании о средиземноморском климате, средиземноморской растительности, животном мире или средиземноморских ландшафтах имеется в виду Средиземноморье именно в этом узком смысле, т. е. Средиземноморский природный регион. Во втором случае к Средиземноморью относятся еще и переходные, или субсредиземноморские, области — север Пиренейского полуострова, Паданская низменность, внутренние районы Балканского полуострова, а также приморская полоса в пределах Ливии и ОАР. Эти области связаны со Средиземноморьем общими этапами истории развития ландшафтов, в их природе проявляются некоторые средиземноморские черты, но тем не менее они уже не являются средиземноморскими в полном смысле этого слова.
Естественно, что территория Средиземноморья не может быть ограничена какими-либо высотными рамками, а должна рассматриваться с присущей ей высотной поясностью, выраженной в горных системах. Она также неотделима от Средиземного моря и морей его бассейна, которые представляют собой неотъемлемый компонент многих средиземноморских ландшафтов.
Тропические Саванны
Тропические саванны (степи с редкими деревьями или группами деревьев) расположены в теплых областях, где в год выпадает большое количество осадков (1000—1500 мм), но имеется один или два продолжительных сухих сезона, когда возникают пожары, представляющие собой важный фактор окружающей среды. Самая обширная область этого типа находится в Центральной и Восточной Африке, но довольно значительные области тропических саванн (grassland) встречаются также в Южной Америке и Австралии. Поскольку и деревья, и злаки должны быть устойчивыми к засухе и пожару, растительность состоит из небольшого числа видов, что составляет разительный контраст с расположенными рядом экваториальными лесами. В покрове доминируют знаки, принадлежащие к таким родам, как Panieum, Pennisetum, Andropogon, Imperata.
Либих (Liebig) Юстус (12.5.1803, Дармштадт, — 18.4.1873, Мюнхен), немецкий химик. Профессор университетов в Гисене (с 1824) и Мюнхене (с 1852). С 1830 член-корреспондент Петербургской АН, с 1860 президент Баварской АН. В 1825 организовал в Гисене лабораторию для практических занятий студентов, в которой учились и работали многие известные химики. Основные исследования в области органической химии. В 1823 нашёл, что гремучекислое серебро AgONC тождественно по составу с циановокислым серебром (первый пример изомерии). В 1832 Либих и Ф. Вёлер показали, что при разнообразных превращениях горькоминдального масла (т. е. бензойного альдегида C6H5COH) неизменно происходит переход из одного соединения в другое одной и той же группы атомов (C6H5CO), названной ими радикалом бензоилом. Работы Л. способствовали утверждению теории радикалов. Впервые (одновременно с французским химиком Э. Субейраном и независимо от него) получил хлороформ (1831) и уксусный альдегид (1835), открыл гиппуровую, молочную и др. карбоновые кислоты. Усовершенствовал методику определения углерода и водорода в органических соединениях (1831—33). С 1839 изучал химизм физиологических процессов, выдвинул химическую теорию брожения и гниения. Л. — один из основателей агрохимии. В 1840 высказал теорию минерального питания растений, способствовавшую широкому внедрению минеральных удобрений в земледелии. Основал (1832) журнал "Annalen der Pharmacie" (в 1840—73 выходил под названием "Annalen der Chemie und Pharmacie", с 1873 переименован в "Liebigs Annalen der Chemie"). Л. создал научную школу, среди его учеников немало видных русских химиков (А. А. Воскресенский, Н. Н. Зинин, Л. Н. Шишков и др.).