егуляторные механизмы СВД.
Постоянство газового состава крови (рО2, рСО2, рН) обеспечивается сложной системой регуляции внешнего дыхания, которая организована как контур отрицательной обратной связи.
Выделяют хеморецепторный контур и механорецепторный контур регуляции.
Хеморецепторный контур представлен двумя основными группами хеморецепторов – это центральные хеморецепторы, ответственные за обнаружение отклонений в гомеостазисе СО2 и каротидные тельца, осуществляющие мониторинг рО2.
Центральные хеморецепторы располагаются вблизи вентральной поверхности продолговатого мозга рядом с нейронами дыхательного центра, ответственными за дыхательный ритмогенез.
Центральные хеморецепторы реагируют на изменения рН и рСО2 крови, причем реакция на рСО2 выражена сильнее, что объясняется легкостью диффузии СО2 через ГЭБ и относительной непроницаемостью для ионов Н+.
Но надо иметь ввиду, что в конечном итоге центральные хеморецепторы реагируют все-таки именно на [H+] во внеклеточной жидкости внуримозгового интерстициального пространства, так как СО2, проникшая через ГЭБ образует угольную кислоту, которая диссоциирует на Н+ и НСО3- .
Каротидные тельца расположены у бифуркации общих сонных артерий. В ответ на артериальную гипоксемию гломусные клетки каротидных телец высвобождают допамин пропорционально степени снижения раО2. Когда раО2 падает ниже 60 мм рт.ст.,активность афферентных нервовкаротидного тельца возрастает экспоненциально, что ведет к формированию гипоксического вентиляционного ответа- повышению вентиляции.Центральные хеморецепторы обеспечивают регуляцию глубины вдоха, а периферические – частоту дыхания.
Механорецепторный контур представлен легочными аорферентными волокнами блуждающего нерва, имеющими 3 группы рецепторов:
1) медленно адаптирующиеся рецепторы растяжения;
2) быстро адаптирующиеся рецепторы растяжения;
3) С –волокна.
1) Медленно адаптирующиеся рецепторы растяжения лежат в гладких мышцах воздухоносных путей и играют важную физиологическую роль в рефлексе Геринга-Брайера, который регулирует окончание вдоха при быстром наполнении легких или, когда дыхательный объем превышает 1 л, и удлинение выдоха.
2) Быстро адаптирующиеся рецепторы – ирритантные рецепторы реагируют на гистамин, лейкотриены, быстрое изменение объема легких, вдыхание пыли, раздражающих газов, холодного воздуха. Стимуляция ирритантных рецепторов вызывает бронхоконстрикцию, частое поверхностное дыхание, кашель и тахикардию.
3) Рецепторы С–волокон оканчиваются в паренхиме легких, ВП и кровеносных сосудах. Они также реагируют на вдыхание раздражающих агентов (газы и частицы) и механическое раздражение воздухоносных путей. Стимуляция С–волокон приводит к:
· частому поверхностному дыханию, которое ограничивает рассеивание вредных агентов по бронхиальному дереву;
· брадикардии;
· секреции слизи и обструкции бронхов;
· ощущению диспное при отеке легких, пневмонии, вдыхании токсических веществ.
В состав С –волокон входят и т.н. юкстакапиллярные или J –рецепторы, которые оканчиваются в паренхиме легких вблизи капилляров. Активация J–рецепторов происходит при отеке легких, пневмонии; вызывает закрытие гортани и апноэ (остановку дыхания) за которым следует частое поверхностное дыхание. J–рецепторы также передают ощущение диспноэ.
Помимо хемо- и механорецепторов поток информации в дыхательный центр обеспечивает ряд других чувствительных структур. Так, стимуляция рецепторов, располагающихся в носовой полости и верхних дыхательных путях приводит к кашлю, чиханию, бронхоконстрикции и спазму гортани.
Проприоцептивные афферентные окончания в периферических суставах и мышцах могут играть существенную роль в усилении вентиляции при физической нагрузке.
Особо надо отметить регуляцию просвета воздухоносных путей, которая осуществляется достаточно автономно, так:
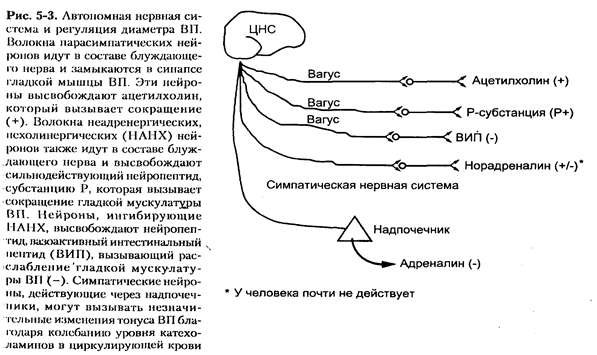
-холинэргические (парасимпатические) метонейроны иннервируют ВП через блуждающий нерв с синапсами, расположенными вблизи от гладких мышц ВП. Эти нейроны высвобождают ацетилхолин, который вызывает сильное сокращение гладких мышц ВП. Эффекты симпатической нервной системы у человека играют незначительную роль в регуляции диаметра ВП.
Сильное влияние на тонус ГМ ВП оказывает т.н. неадренергическая, нехолинергическая (НАНХ) нервная система. Одна группа нейронов НАНХ системы включена в локальные рефлекторные дуги (аксон-рефлексы), в которых местное раздражение нервных окончаний, при повреждении эпитениальных клеток или под действием медиаторов воспаления, ведет к высвобождению субстанции Р и сокращению гладких мышц ВП. Другая группа нейронов НАНХ системы высвобождает активный интестинальный пептид (ВИП), который расслабляет гладкие мышцы ВП посредством повышения внутриклеточного цАМФ.