Особливості біології бурих водоростей.
Зміст
Вступ
Розмноження водоростей
Особливості біології бурих водоростей
Загальні ознаки вищих рослин
Основні форми талломів водоростей
Різноманітність бурих водоростей
Значення водоростей у природі та житті людини
Висновок
Список використаної літератури
Вступ
Водорості — велика група найдавніших рослин. Будова їхнього тіла і розміри характеризуються значною різноманітністю. Існують одноклітинні, багатоклітинні і колоніальні форми мікроскопічних розмірів (від тисячних часток міліметра), а також форми з різною будовою слані, що досягають 30—45 м.Водорості — єдина група організмів, серед яких зустрічаються прокаріоти (синьозелені) і еукаріоти (решта відділів). В ядрах еукаріотних водоростей виявлені структури, властиві ядрам інших еукаріотів: оболонки, ядерний сік, ядерця, хромосоми.
Загальною ознакою всіх водоростей є наявність хлорофілу. Крім хлорофілу водорості можуть містити й інші пігменти (фікоціан, фікоеритрин, каротин, ксантофіл, фіко-сантин). Щ пігменти надають водоростям червоного, бурого, жовто-зеленого кольору, маскуючи основний зелений. Наявність пігментів у клітинах водоростей забезпечує автотрофний тип живлення. Проте багато водоростей здатні за певних умов переходити на гетеротрофне живлення (евгленові — в темряві) або поєднувати його з фотосинтезом (міксотрофний тип живлення).
Кількість видів водоростей перевищує 40 тис. Проте класифікація їх не завершена, оскільки не всі форми достатньо вивчені. У нашій країні прийнято поділяти водорості на 10 відділів: синьозелені, пірофітові, золотисті, діатомові, жовтозелені, бурі, червоні, евгленові, зелені, харові. Найбільшу кількість видів налічують зелені (13—20 тис.) і діатомові (10 тис.) водорості.
Поділ водоростей на відділи збігається зазвичай з їхнім забарвленням, яке, як правило, пов'язане з особливостями будови клітин і слані.
Будова, склад і властивості клітинних компонентів водоростей характеризуються великою різноманітністю. У процесі еволюції природний добір зберіг найперспек-тивніші форми, в тому числі такий тип клітинної організації, який дав змогу рослинам перейти до наземного способу життя.
Розмноження водоростей буває вегетативним, безстатевим (за допомогою спор) і статевим. У одного й того самого виду залежно від умов і пори року способи розмноження різні. При цьому спостерігається зміна ядерних фаз — гаплоїдної і диплоїдної.
Сприятливими умовами для зростання водоростей є наявність світла, джерел вуглецю та мінеральних солей, а основним середовищем життя для них є вода.Значний вплив на життя водоростей мають температура, солоність води тощо.
За місцем зростання водорості поділяють на дві великі групи: ті, що живуть у водоймах, та ті, що живуть поза водоймами. Серед тих, що живуть у водоймах, виділяють такі екологічні групи: планктон — дуже дрібні водорості, які знаходяться в товщі води у завислому стані (хламідомонада, вольвокс, пандорина, мікроцистіс); бентос — водорості, що живуть на дні водойм (харові, багато діатомових); перифітон — водорості, якими обростають підводні предмети або вищі рослини водойм (кладофора, червоні та бурі водорості морів). Населяють водорості і гарячі джерела, а також водойми з підвищеною концентрацією солей.
Водорості, що живуть поза водоймами, також поділяють на групи: грунтові (едафітон) — живуть у грунті або на грунті (ботридій, деякі вошерії), у грунті живе понад
700 видів водоростей з різних відділів; наземні (аерофі-тон) — на корі дерев, на скелях (трентеполія, плеврокок).
Багато водоростей вступають у симбіотичні зв'язки з іншими представниками рослинного й тваринного світу. Особливе місце тут належить симбіозу водоростей з грибами. У цьому симбіозі виникає така біологічна єдність двох організмів, що призводить до появи третього — ли-шайника, який відрізняється і від першого, і від другого.
Зелені водорості. Ознайомимося з представниками відділу зелених водоростей, розглянувши з одноклітинних хламідомонаду, плеврокок і хлорелу, з багатоклітинних нитчастих — улотрикс і спірогіру.
Хламідомонада — мікроскопічна одноклітинна водорість грушоподібної або овальної форми. Поверхня клітини вкрита прозорою безбарвною пектиновою оболонкою. На передньому кінці тіла оболонка утворює невелике випинання — носик, від якого відходять два джгутики. За допомогою цих джгутиків водорість рухається. Всю внутрішню частину клітини займає цитоплазма з великим чашоподібним хлоропластом (хроматофором) зеленого кольору. В нижній потовщеній частині хлоропласта розміщений кулястий піреноїд (зона, де найактивніше синтезуються й нагромаджуються поживні речовини), який містить багато білків і оточений зернами крохмалю. На верхній частині хроматофора знаходиться добре помітне червоне вічко, насичене каротином. Функція його невідома. В заглибині хроматофора розміщене велике кулясте ядро з добре помітним ядерцем. Спереду біля основи джгутиків розміщені дві пульсівні вакуолі.
Хламідомонади розмножуються статевим і безстатевим шляхом .При безстатевому розмноженні клітина втрачає джгутики, її ядро, хлоропласт і цитоплазма Діляться на 4 (рідше 8) клітин — зооспор. У кожної дочірньої клітини виростає по 2 джгутики, оболонка материнської клітини руйнується і зооспори виходять у воду.
У такий спосіб водорості розмножуються дуже швидко. Же через добу дочірні клітини знов діляться. При статевому розмноженні в материнській клітині утворюються гамети. Вони подібні до зооспор, однак
кількість значно більша — 32 або 64 в одній клітині. Після дозрівання гамети виходять з материнської клітини і попарно з'єднуються, утворюючи зиготу. Вона вкривається исною оболонкою і переходить у стан спокою. Через який час зигота виходить з оболонки і ділиться мейочно з утворенням 4 гаплоїдних зооспор.

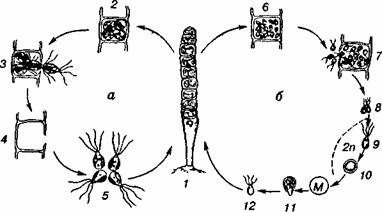
Мал.1 Загальна характеристика нижчих рослин – водоростей. Особливості біології зелених водоростей.
. Життєвий цикл хламідомонади:
а — безстатеве розмноження; б — статеве розмноження; М ~ мейоз; 1 — вегетативна особина; 2 — утворення зооспор; 3 — зооспори; 4 — утворення гамет; 5 — копуляція гамет; 6 — зигота; 7 — проростання зиготи; 8 — зооспора
Хламідомонади живуть у невеликих, дуже забруднених водоймах, що добре прогріваються (калюжі, стічні канави). У таких водоймах вони дуже швидко розмножуються і спричинюють "цвітіння" води. Оскільки поряд з автотрофним способом живлення клітини хламідомонад поглинають розчинені органічні речовини, вони тим самим сприяють процесу очищення забрудненої води (самоочищенню). Цей процес можна спостерігати у відстійниках, очисних спорудах міської каналізації.
Уже через кілька днів після чергового викидання стічних вод водойма очищається. Вода стає чистою і прозорою до дна. Водоростей — активних санітарів — уже немає в товщі води, їхні зиготи опустились на дно і там "відпочивають" до чергового забруднення, яке знов їх активізує.
Плеврокок — мікроскопічна одноклітинна наземна водорість без джгутиків. Під щільною безбарвною оболонкою клітини знаходяться цитоплазма, ядро і пластинчастий хлоропласт. Зазвичай клітини поєднані в групи по 4—6 і більше у вигляді пакетів, іноді утворюють короткі нитки. Пакети можуть розпадатися на окремі клітини» кожна з них існує самостійно і швидко починає ділитися.
Клітини плеврокока діляться в двох взаємно перпендикулярних напрямках, утворюючи нові пакети. Статевого розмноження немає, зооспори не утворюються. Поселяється плеврокок на корі старих дерев, особливо на північному боці на землі в сирих місцях, на стінках глиняних горщиків з кімнатними рослинами, утворюючи на них разом з іншими водоростями зелений наліт. Плеврокок входить до складу слані багатьох лишайників.
Хлорела — одноклітинна зелена водорість, має вигляд мікроскопічної нерухомої (без джгутиків) кульки до. 15 мкм у діаметрі. Зовні клітини вкриті твердою двоконтурною оболонкою целюлозної природи. В цитоплазмі міститься один чашоподібний хлоропласт з одним піре-ноїдом у потовщеній його частині. Ядро одне, однак у живій клітині без спеціальної обробки його не видно.
Розмножується лише нестатевим шляхом. При цьому в материнській клітині утворюється 4—8 апланоспор, які через розрив оболонки виходять у воду й набувають вигляду дорослої особини.
Хлорела невибаглива до умов існування і здатна до інтенсивного розмноження, тому зустрічається всюди: у прісних водоймах, морях і грунті. Вона може вступати у симбіоз з різними організмами, входить до складу лишайників.
Клітина хлорели — зручний об'єкт для різних досліджень. Хлорела — основний об'єкт масового культивування водоростей для практичного використання в різних напрямах. Значну роль у формуванні підвищеного інтересу до неї відіграв її хімічний склад. У перерахунку на суху речовину хлорела містить повноцінних білків 40 % і більше, ліпідів — до 20 %, вуглеводів — до 35 %, зольних речовин — до 10 %. Є вітаміни групи В, аскорбінова кислота (віт. С) і філохінони (віт. К). Знайдено речовину, яка має антибіотичну активність — "хлорелін". У деяких країнах хлорелу використовують у їжу після спеціальної обробки, Що поліпшує її засвоєння. Для споживання використовують свіжу біомасу хлорели або спеціальну пасту з неї.
Крім одноклітинних форм до відділу зелених водоростей належать і багатоклітинні нитчасті. Форма їхнього тіла має вигляд простих або розгалужених ниток, що безперервно ростуть у довжину в результаті поділу клітин упоперек. Найтиповішими представниками нитчастих водоростей є улотрикс і спірогіра.
Улотрикс дуже поширений у річках, живе, прикріплюючись до підводних предметів, утворюючи яскраво-зелені ростання. Баговиння улотрикса складається з нерозгалужених ниток різної довжини, які на початку росту прикріплюються до субстрату безбарвною видовженою клітиною — ризоїдом. Клітини ниток циліндричні або бочкоподібні, короткі. Кожна клітина має ядро, пристінний хлоропласт у вигляді неповного кільця і один або кілька піреноїдів.
Розмноження
Розмноження вегетативне, безстатеве і статеве (мал. 2). При вегетативному розмноженні нитка улотрикса розпадається на короткі сегменти, кожний з яких росте, утворюючи нову нитку.
Безстатеве розмноження здійснюється за допомогою зооспор, які формуються в клітинах. Зооспори — яйце-або грушоподібні клітини з чотирма джгутиками на передньому кінці, червоним вічком і двома пульсівними вакуолями. Кожна зооспора, вийшовши назовні, через деякий час прикріплюється до субстрату і проростає в дорослу особину.
При статевому розмноженні в клітинах нитки формуються гамети, подібні до зооспор, однак лише з двома джгутиками. Гамет у кожній клітині більше, ніж зооспор. З'єднуючись попарно, гамети однієї й тієї самої або частіше різних ниток утворюють зиготу. Вона залишається рухливою недовго, скоро осідає на дно водойми, втрачає джгутики і вкривається товстою оболонкою, що захищає ЇЇ від несприятливих умов. Під час проростання зиготи відбувається редукційний поділ з утворенням 4—16 гаплоїдних особин.
Мал. 2. Життєвий цикл улотрикса:
а — безстатеве розмноження; б — статеве розмноження; М — мейоз; / — вегетативна особина; 2 — утворення зооспор; 3 — вихід зооспор; 4 — порожня клітина; 5 — зооспори; 6 — утворення гамет; 7 — вихід гамет; 8 — ізогамія; 9, 10 — зиготи; 11 — проростання зиготи; 12 — зооспора
Спірогіра — одна з найпоширеніших зелених нитчастих водоростей у прісноводних басейнах. Довгі нитки слані утворюють сплетення (баговиння) яскраво-зеленого кольору, до субстрату вони не прикріплюються і вільно плавають у воді.
Нитки спірогіри завдовжки від кількох міліметрів до 8-10 см не галузяться і складаються з одного ряду однакових видовжених циліндричних клітин. Кожна клітина має двошарову оболонку, яка зовні вкрита слизовим чохлом. Внутрішній шар оболонки складається з целюлози, зовнішній — з пектинових речовин. Целюлозна оболонка оточує цитоплазму, в якій розміщені спіральне закручені зелені стрічкоподібні хлоропласти з численними піреноїдами. Велике ядро з добре помітним ядерцем розміщене в центральній частині клітини, оточене шаром цитоплазми, який сполучений з пристінним шаром тяжами. Проміжки між тяжами заповнені вакуолями.
Спірогіра розмножується вегетативне і статевим шляхом, спор вона ніколи не утворює. Вегетативне розмноження відбувається в разі випадкового розриву ниток або розпадання її на окремі клітини за несприятливих умов. З кожної частини нитки або окремої клітини утворюються нові особини поділом клітин.
Статеве розмноження здійснюється кон'югацією. При цьому дві нитки, зовні подібні, зближуються. З клітин ниток, розміщених одна навпроти одної, утворюються вирости. При стиканні цих виростів (мал. 3) перегородки їх зникають і вміст "чоловічої" клітини переливається в "жіночу". В результаті цього злиття утворюється зигота, яка вкривається товстою оболонкою і після певного періоду спокою проростає. При цьому вона ділиться мейозом, утворюючи чотири гаплоїдні клітини. Три з них — дрібні — відмирають, а одна — велика — разом із вмістом зиготи утворює проросток нової особини, який дає початок новій нитці.
У ряді випадків з'ясовані (експериментально) умови зовнішнього середовища, що стимулюють перехід до зооспорообразованію. Для багатьох нитчастих зелених водоростей таким чинником служить підвищений вміст у воді вільної вуглекислоти. Це пояснює старі спостереження над масовою зооспорообразованіем при перенесенні ниток із текучої води в стоячу, зі світла в темряву, тобто в умови, коли у воді накопичується вуглекислота.
У зв'язку з зооспорообразованіем доречно зупинитися на виникненні так званих сінзооспор. Сінзооспори - освіти, відповідні комплексу двох чи багатьох зрощених в різного ступеня зооспор, - спостерігалися у різних водоростей, але особливо часто у разножгутікових. Нерідко при виході з материнської клітини такі зооспори залишаються з'єднаними по дві або по декілька лише за допомогою тонких цитоплазматичних ниток і пізніше роз'єднуються. Однак часто зооспори виявляються пов'язаними своїми задніми кінцями набагато міцніше і на більшому протязі, так що вільними залишаються тільки їх передні кінці з джгутиками або ж, нарешті, вони утворюють суцільну цитоплазматичну масу, з якої виступають пари джгутиків, вказуючи на число об'єдналися зооспор. Такі стадії дуже схожі на сінзооспори вошерії, яким вони повністю відповідають.
Сінзооспори часто зустрічаються у бочрідіопсіса. трібонеми, ботрідіума та ін, однак, як правило, ці водорості утворюють двожгутикові зооспори. Ці випадкові відхилення стабілізувалися лише у вошерії. Її великі многожгутіковие зооспори вже давно розглядали як складну зооспор, що утворюється всередині спорангия, в якому поділ спорапгіального протопласта на одноядерні двужгутіковие суперечки не спостерігається.
Сінзооспори є також в зелених (сфероплея, кладофора, хламідомонас тощо) і бурих водоростей (Ectocarpus siliculosus).
У значної кількості водоростей замість, зооспор утворюються цпланрслори - нерухомі, позбавлені джгутиків суперечки. Прикладами їх можуть служити моно-і тетраспори ктеасних водоростей, тетраспори діктіотових з бурих водоростей. У тих випадках, коли апланоспори, будучи в'язнями в оболонку материнської клітини, беруть усі відмітні риси цієї клітини (характерні обриси, особливості оболонки), їх називають автоспорамі (наприклад, у багатьох хлорококкальних із зелених водоростей).
У ценобіальних (монадне і коккоідних) зелених водоростей при безстатевому розмноженні утворюються дочірні колонії. У ряді груп водоростей безстатеве розмноження відсутній: у кон'югатофіціевих, харофіціевих, багатьох зелених водоростей сифонові будови, у всіх діатомових, фукальних з бурих водоростей.
На прикладі деяких водоростей вдалося глибше проникнути в механізми звільнення кліток безстатевого розмноження та їх подальшого прикріплення до субстрату. Ось деякі приклади. Для багатьох (понад 40) видів хламідомонади було показано, що молоді клітини виділяють літичний фактор - автолізін спорангіальной стінки, викликає лізис клітинної стінки спорангия, що сприяє звільненню зооспор. Фермент діє тільки на спорангіальную стінку, але не на стінки вегетативних клітин і зооспор і володіє груповий специфічністю, тобто: чи впливає тільки на морфологічні подібні види, а в деяких випадках - тільки на спорангії виду-продуцента.
У вольвоксу клітини також утворюють і виділяють у навколишне середовище ферменти, лизирующие інволюкрум материнського ценобія. полегшуючи цим звільнення дочірніх сфероїд. З культурального середовища Chiorella fusca var. vacuolates був виділений фермент (відсутній в середовищі до початку сноруляціі), руйнує клітинну стінку материнських клітин автоспор і не діє на клітинні стінки самих автоспор, У вошерії ст. кінчику сінзооспори були виявлені численні везикули, мабуть, деривати діктіосом, що містять імовірно целюлазу та інші ензими, що руйнують полісахариди. Висловлено припущення, що вони відповідальні за руйнування стінки в апікальній області спорангия - необхідна умова виходу сінзооспори. У зооспор зеленої водорості Oedogonium cardiacum, бурої - Chorda tomentosa, тетраспорах червоної водорості Palmaria palmata виявлені рясні везикули, що містять клеючий матеріал, який, можливо, сприяє прикріпленню спір до субстрату. Те ж, мабуть, справедливо і для тетраспор і карпоспор Ceramium rubrum.
Статеве розмноження (відтворення), сутність якого полягає у злитті - копуляції двох різних в статевому відношенні клітин-гамет з утворенням зиготи, відомо у більшості еукаріотичних водоростей. У багатьох випадках, особливо у водоростей з монадної організацію, виявленню статевого процесу сприяла розробка методів їх культивування в лабораторних умовах. Наприклад, при певних умовах культивування в останні роки був описаний статевий процес у ряду дінофлагеллят, що раніше вважалися позбавленими його.
Статеве розмноження - складний багатоступінчастий процес, який з статевого диференціювання клітин-гаметогенезу, що включає злиття гамет (те, що відбувається в кілька етапів і приводить до утворення зиготи) і завершується мейозом.
Умови, що викликають статевий процес. Перехід до статевого розмноження і диференціювання гамет залежить не тільки від внутрішніх причин, нерідко обумовлених віком водорості, але і від факторів зовнішнього середовища. Серед цих факторів для багатьох зелених водоростей провідну роль відіграє зміст в середовищі азоту. Вичерпання в середовищі азоту і наступаюче азотисте голодування служить сигналом для початку статевого розмноження багатьох видів Chlamydomonas (Ch. reinhardtii, Ch. Moewusii var. Rotunda, Ch. Chlamydogama, Ch. Eugametos), Pandorina morum, P. unicocca, Chlorococcum echinozygotum, Golenkinia minutissima, Scenedesmus obliquus, Oedogonium cardiacum та ін Хорошим прикладом залежності статевого диференціювання від вмісту в середовищі азоту служить зелена водорість Sphaeroplea annulina. Ця азотолюбівая водорість зустрічається у нас переважно ранньою весною в дрібних водоймищах і вже в травні при зниженні вмісту у воді азоту переходить до статевого відтворення. У водоймах з більш постійним підтікання азотистих речовин, наприклад на полях зрошення Москви, сфероплея вегетирует іноді майже все літо. У лабораторії також можна підтримати її у вегетативному стані, культивуючи в середовищі зі значним вмістом азоту; перенесенням ж у середу, бідну солями азоту, легко можна викликати статевий процес.
У перелічених та багатьох інших видів джерела азоту (нітрати, амоній, сечовина, глютамін та ін), що підтримують зростання, перешкоджають статевої диференціювання, яка наступає в умовах, несприятливих для вегетативного росту. Ріст і розмноження перебувають ніби в антагоністичних відносинах один з одним. На відміну від більшості зелених водоростей у вольвокс статеве розмноження з подальшим утворенням покояться зигот відбувається в той час, коли умови середовища оптимальні для вегетативного росту. Це пояснюється тим, що статевий процес у цього роду пов'язаний з продукцією спеціальних статевих ценобіев, в яких і формуються гамети. У менш оптимальних для вегетативного росту умовах статеві сфероїд зменшувалися б у розмірах, знижувалася б їх вітальність. Статева диференціювання у вольвокс настає під дією видоспеціфічних статевих гормонів - індукторів, що виділяються чоловічими ценобіямі, що з'являються в популяції спонтанно.
Дефіцит в середовищі азоту викликає статевий процес і у багатьох дінофлагеллят.
Для індукування статевого процесу у десмідіальних водоростей найбільш важливими факторами виявляються світло і температура, хоча в деяких випадках формування кон'югаційні виростів і освіта зигот спостерігалося при нестачі в середовищі азоту. Світло і температура є також вирішальними чинниками середовища, що надають вплив на гаметогенез багатьох бурих водоростей. Так, для багатьох ламінаріальних водоростей було показано, що перехід їх гаметофитов до гаметогенезу контролюється синім світлом досить високої інтенсивності при низьких (5-10 ° С) температурах. У організмів, приурочених до вод з високим вмістом солей, наприклад у Dunaliella salina, зниження концентрації солей у середовищі викликає статевий процес.
Форми статевого процесу. Форма статевого процесу може бути різною. У низки позбавлених клітинної оболонки монадне водоростей зливаються, (копуліруют) клітини, не відрізняються від вегетативних. Така форма статевого процесу називається хологамия (наприклад, у зеленій водорості дуналієлла). У багатьох водоростей гамети, як і зооспори, утворюються всередині материнських клітин і, вийшовши з них, копуліруют попарно. Якщо обидві зливаються монадне гамети не розрізняються морфологічно, статевий процес називають изогамного - изогамия, при каплиці монадне гамет, різних за розміром, має місце гетерогамия або анізогамія. Більші макрогамети трактуються як жіночі, більш дрібні - мікрогамети - як чоловічі. При моогамном статевому процесі - великі жіночі гамети позбавлені шулшов, нерухомі і називаються яйцеклітинами або з яйцями, більш дрібні чоловічі гамети з джгутиками рухливі і носять назву сперматозоїдів або антерозоідов.
У червоних водоростей яйцеклітини запліднюються чоловічими клітинами, позбавленими джгутиків, - спермаціями.
У зелених водоростей за поодинокими винятками спостерігається справжня оогамия, для якої характерно запліднення яйця всередині оогоній - на місці утворення (in situ) на відміну від примітивної оогамия, коли яйцеклітина покидає оогонії до запліднення, що здійснюється в навколишньому середовищі. Примітивна оогамия зустрічається серед зелених водоростей лише у вигляді винятків, є правилом для оогампих бурих водоростей. У центрофіціевих оогамном діатомових спостерігається як справжня, так і примітивна оогамия. Якщо зливаються протопласти нерухомих клітин, що не відрізняються від вегетативних, говорять про кон'югації.
Звичайно передбачається, що форма статевого процесу еволюціонувала в наступному напрямку: изогамия-»анізогамія-» оогамия. Ця прогресивна еволюція почалася з виникнення різних за розміром гамет у изогамного предкової форм. Освіта дрібних гамет підвищує їх нумерично продуктивність - їх число, але за рахунок редукції кількості цитоплазми. З іншого боку, велика маса цитоплазми і резервних матеріалів у більш великих гамет (що виникають в материнських клітинах у меншій кількості) необхідні для початкового розвитку майбутньої зиготи. Таким чином, раз виник диморфізм гамет дає основу для виборчого злиття між гаметами, які відрізняються за розмірами: гамети мінімальних розмірів отримують компенсацію у кількості цитоплазми допомогою злиття з більшими гаметами. Під ці суто теоретичні міркування останнім часом була підведена експериментальна основа.
Л. Візе вдалося експериментально отримати фенотипическую анізогамію у трьох изогамного гетероталлічних видів Chlamydomonas. У цих видів кожен тип спаровування («+» і «-») має два шляхи гаметогенезу, які індукуються підбором умов культивування. Перший шлях гаметогенезу полягає в тому, що протопласт вегетативної клітини в результаті внутрішньоклітинної перебудови безпосередньо перетворюється в одну велику гамету. При другому способі гаметогенезу в материнській клітині відбуваються два або три мітозу, і утворюється 4-8 гамет меншого розміру. Комбінуванням сумісних гамет, що сформувалися різними способами, може бути отримана фенотипічно анизогамного копуляція. Висловлено припущення, що будь-яка мутація, яка зафіксувала б той чи інший спосіб гаметогенезу (блокувала б один шлях гаметогенезу на користь іншого), дала б безпосередньо штами, що виробляють виключно або мікро-, або макрогамети. Таким чином, могли виникнути вже генетично анизогамного лінії. У який з'явився в результаті мутації анизогамного таксонів при подальшої еволюції може бути втрачена рухливість у більш великих гамет.
Еволюція форми статевого процесу відбувалася незалежно від еволюції таллома. Це вперше продемонстрував російський альголог І.М. Горожанкін в кінці минулого століття, виявивши у різних видів хламідомонади (що стоять на одному рівні морфологічної диференціювання таллома) всі основні типи статевого процесу: поряд з изогамия, переважаючою в одноклітинних вольвокальних, гетерогамия у Ch. braunii і оогамия у Ch. coccifera.
Гаметогенез. У форм з изогамного і гетерогамним статевим процесом гаметогенез в більшості випадків не пов'язаний з формуванням будь-яких відокремлених морфологічних структур: з-і гетерогамети виникають у звичайних вегетативних клітинах, як і зооспори, але звичайно в більшому числі і відповідно менших розмірів. Виняток становлять изогамного і гетерогамние бурі водорості, де гамети утворюються в спеціальних органах: багатогніздна або багатокамерних спорангіях, в кожній камері (клітині) яких формується за однією гаметі. Оогамия у монадне і коккоідних форм зазвичай також не пов'язана зі зміною клітин, що виробляють гамети: вони не відрізняються від вегетативних. Навпаки, у водоростей з нитчатой і тканинної організацією таллома яйцеклітини і сперматозоїди розвиваються в гаметангиях, як правило, різко відрізняються за формою від вегетативних клітин і носять спеціальні назви оогонії і антеридії (сперматогонії).
Рослини, що виробляють гамети, можуть бути двостатевими (однодомних) і роздільностатеві (дводомні). У першому випадку гамети різної статі виникають на одному і тому ж таллома, у другому - на різних. Крім понять «однодомна» і «дводомність» існують поняття «гомоталлізм» і «гетероталлізм». При гомоталлізме статевий процес має місце в межах клональною популяції (клон - культура, отримана з однієї клітини). При гетероталлізме статевий процес можливий тільки при змішуванні клітин з різних клонів протилежного типу спаровування або знака; у разі гетероталлічних изогамного форм гамети різного типу спарювання позначаються знаками «+» і «-». Поняття «гомоталлічний» і «гетероталлічний» не завжди збігаються з поняттями «однодомних» і «дводомний». Так, серед вольвокс є гомоталлічние і гетероталлічние види, що включають як однодомні, так і дводомні штами. Наприклад, всередині клональною популяції можуть розвиватися особини як однодомні з яйцями і сперматозоїдами в одному сфероїд, так і дводомні, у яких статеві органи перебувають у різних індивідах.
Звільнення репродуктивних клітин. Деякі морські водорості, що живуть у верхній частині літоральної зони, виявляють періодичність у формуванні та звільнення репродуктивних клітин як безстатевого (суперечка), так і статевого (гамет) розмноження. Так, біля берегів Англії у водорості Dictyota dichotoma формування статевих органів (антеридиев і оогоній) приурочене до літніх місяців і відбувається періодично з двотижневими інтервалами, що відповідає таким між двома сігізійнимі припливами, а звільнення гамет - яєць і сперматозоїдів - настає через кілька припливів після найвищого з наступних квадратурних припливів і триває 7-9 днів. Таким чином, періодичність знаходиться в строгій відповідності з місячним ритмом. Періодичність зберігається і при культивуванні талломов в лабораторії. У одного й того ж виду ритм звільнення гамет залежить від географічного положення місцеперебування. Так, у D. dichotoma в Північній Кароліні періодичність спостерігалася, але інша, ніж у берегів Англії: гамети звільнялися не через двотижневі інтервали, а через місячні, а біля берегів Ямайки статеві органи формувалися періодично, але період дозрівання гамет розтягувався настільки, що вихід їх відбувався майже безперервно.
Поряд з напівмісячним ритмом у звільненні яйцеклітин у D. dichotoma була описана і денна періодичність. При культивуванні в умовах зміни світла (14 год) і темряви (10 год) незабаром після початку світлового періоду спостерігався активний вихід яйцеклітин, який тривав протягом години, після чого звільнення яєць припинялося до початку наступного світлового періоду, коли знову починалося звільнення яєць. Денний вихід яєць може бути індукований дуже слабким короткочасним освітленням. Так, 50% яєць зрілого оогоній звільняється після 20-секундного освітлення білим світлом в 1 Лк. Вихід яєць залежав також від температури: якщо при 20С максимум звільнення яєць досягається через 25-30 хв після початку денного світлового періоду, то при 10С цей максимум наступав лише через 100 хв. Ефект світла даній інтенсивності також залежить від температури. Крім того, розкриття повністю дозрілого оогоній може бути прискорене або уповільнити шляхом зміни осмотичного тиску середовища.
Спеціальне дослідження щодо з'ясування впливу деяких зовнішніх умов на періодичність звільнення гаморі було проведено в Японії над популяційними звичайної в прибережних водах водорості - Monostroma nitidum, що займає друге (після видів порфіри) місце в якості об'єкту промислового культивування. Це культивування здійснюється зазвичай двома методами: на хібісетках, розташованих горизонтально на такій глибині, при якій талломи оголюються під час відливу приблизно на 4 години, і на бета-сітках з рослинами, постійно зануреними в морську воду поблизу її поверхні. Освіта гамет легко розпізнається неозброєним оком завдяки жовтуватою забарвленні плодущіх частини таллома. Були проведені порівняльні спостереження над таллома, виростають поблизу один від одного, але в розрізняються умовах: на прибережних скелях - в природних місцях проживання, на хібі-сітках і бета-сітках. Період плодоношення (з лютого по червень) в усіх гріх випадках був подібним, а періодичність у звільненні гамет виявилася різною. Більш чітко (з двотижневими інтервалами, під час кожного квадратурного припливу) періодичність була виражена у рослин, що ростуть на скелях. Менш чітко вона виявлялася у талломов на хібі-сітках, а в постійно занурених талломов на бета-сітках ніякої періодичності не спостерігалося. Продемонстровано також, що періодичність в освіті гамет залежить від освітлення (інтенсивність світла і тривалість фотоперіоду) і температури.
Періодичність спостерігалася і в спороношення Viva pertusa, у звільненні яйцеклітин у Sargassum muticum.
В останні роки в ряді робіт простежено диференціювання і механізм звільнення спермациями у червоних водоростей на електронно-мікроскопічному рівні. Так, у Ptilota plumosa спер-матогоніі, в яких утворюються чоловічі гамети - сперманіі, мають чітко висловленої полярністю, що полягає в тому, що клітинне ядро займає апікальне положення, а в базальної частини молодого сперматогоіія знаходиться одна або декілька вакуоль. У зрілих сперматогонії вакуолі заміщуються великими фіброзними покресленими вакуолями, які займають приблизно половину об'єму клітини і що виникають, можливо, за рахунок злиття пухирців, що походять від діктіосом. Фіброзні вакуолі містять мукополісахаридом, Навколо клітинного ядра і навколишнього цитоплазми, мабуть, утворюється нова плазмалемма, що не включає базальних фіброзних смугастих вакуоль. Таким чином формується спермациями, який залишає оболонку сперматогонії. Одночасно з розривом клітинної стінки сперматогонії спорожняються базальні великі фіброзні вакуолі, що сприяє звільненню спермациями. Спермациями без клітинної оболонки одягнений тонкої слизової обгорткою, можливо походить від матриксу фіброзної вакуолі. Ця обгортка грає важливу роль під час прилипання спермациями до трихогину. Схожі процеси спостерігаються і при диференціювання і звільнення спермациями у Polysiphonia hendryi. У ламінарії при утворенні сперматозоїдів апарат Гольджівиділяє велику кількість слизу, сприяє розриву антерідію і звільненню сперматозоїдів.
У вошерії везикули, можливо, містять целюлази, спостерігалися в апікальній частині оогоніального дзьобика, у місця, де сперматозоїди входять в оогоній незабаром після того, як дзьобик утворює пору. Такі ж везикули спостерігалися поблизу кінчика антерідію, де виходять сперматозоїди.
Копуляція. Злиття - спаровування гамет, що приводить до виникнення зигот, відбувається в кілька етапів. Перший з цих етапів - утворення груп, в яких гамети різної статі вступають в первинний контакт. Якщо у краплі води або культуральної рідини спостерігати поведінку забезпечених джгутиками гамет однієї статі (у разі ізогамет - одного типу спаровування або одного статевого знаку «+» або «-»), можна переконатися, що гамети рівномірно розсіяні по всьому полю зору мікроскопа. Варто до краплі з такими гаметами додати гамети протилежної статі (знаку), як зараз же утворюються скупчення - групи. Це вперше спостерігалося наприкінці минулого століття у изогамного бурої водорості Ectocarpux siliculosus, де багато гамет однієї статі оточують одну гамету протилежної статі. Це було протягнуто моістріровано із застосуванням вітального фарбування гамет протилежних статей у різні кольори. Подібним чином утворюються групи гамет у Chlamydomonas раірега.
В інших водоростей кожна група містить по багато гамет обох статей, що показано у Dunalielld saltna шляхом змішування пофарбованих у зелений колір гамет однієї статі (що утворилися в культурі з надлишком азоту і фосфору) з пофарбованими в помаранчевий колір гаметами протилежної статі (така фарбування виникає в культурах з нестачею в середовищі азоту і фосфору). Серед анизогамного водоростей, наприклад у Ulva lobata,мікроскопічні спостереження показують, що в одній групі є багато гамет обох розмірів.
Освіта груп вже давно пов'язували з виділенням якихось статевих речовин (атрактантів), хемотаксичних залучають гамети протилежної статі. На такий висновок наштовхували і самі ранні спостереження за статевим процесом у водоростей (фукуса, едогониум).
У ряді випадків присутність залучають статевих речовин було продемонстровано експериментально, наприклад, у зеленій водорості Sphueroplea annulina. У неї спостерігається оогамном статевий процес: в одних клітинах (оогоніальних) утворюється багато яйцеклітин, в інших (антерідіальних) - колосальна кількість двужгутікових сперматозоїдів золотистого кольору. При дозріванні статевих клітин у стінках оогонії і антеридиев виникають отвори. Що вийшли сперматозоїди скупчуються біля отворів, що ведуть в оогоній, вриваються в оогоній і своїми рухами приводять у рух яйцеклітини. Якщо серед ниток сфероплеі із зрілими оогоніями помістити бавовняні ниточки, вони абсорбують передбачуваний агтрактант і, перенесені в рівномірну суспензію сперматозоїдів, залучають їх: сперматозоїди скупчуються навколо таких ниточок. Виділення атрактантів гаметами Tetraspora lubrica було продемонстровано іншим способом: при додаванні в рідину з-під гамет одного знака, віддалених центрифугуванням, гамет протилежного знаку, останні утворювали групи. Така ізоаглютінація спостерігається і в інших водоростей.
Робилися також спроби встановити хімічну природу атрактантів (інакше Гамоні). Однак все більш ранні спроби виявилися марними, що значною мірою пояснюється обмеженими в той час можливостями аналітичної хімії. Лити у 70-ті роки XX ст. для декількох морських бурих водоростей вдалося встановити точну хімічну природу атрактантів. Це стало можливим тільки завдяки розвитку методів аналітичної хімії, а також у зв'язку з можливістю культивування водоростей в умовах лабораторії, що дозволяє накопичити більшу біомасу, необхідну для біохімічних досліджень.
У деяких водоростей атрактанти, мабуть, відсутні. Наприклад, у Volvox rousseletii глобоіди сперматозоїдів, які звільнилися з чоловічих ценобіев. плавають у воді швидко, і в присутності жіночих ценобіев можуть прикріпитися до них. Однак специфічне залучення відсутня, скоріше, має місце випадкове зіткнення. Нерідко спостерігаються глобоіди сперматозоїдів, прикріплені до вегетативних і чоловічим сфероїд.
У групах здійснюється первинний контакт гамет та їх попарне злиття, що починається зі злиття їх цитоплазми - плазмогаміі.
Використання альгологіческі чистих і аксенічних культур водоростей і розвиток методів, що дозволяють індукувати гаметогенез, зробили можливим більш ретельне і детальне дослідження процесів запліднення у водоростей як у світловому, так і в електронному мікроскопі, а іноді навіть із застосуванням мікрокіпематографіі (наприклад, у Prasiola scipitata, AscophyUum nodosum, Halidrys siliquosa, Oedogonium cardiacum). У ряді випадків процес дійсного злиття статевих клітин вдалося спостерігати при використанні штамів, спеціально отриманих в умовах культури і не зустрічаються в природі. Так, злиття сперматозоїда з яйцеклітиною у вольвоксу (Volvox carteri f. nagariensis) вдалося спостерігати у жіночого мутанта, що характеризується великим числом яєць і невеликим (порівняно з диким штамом) кількістю соматичних клітин - велика кількість їх ускладнювало спостереження цього процесу. У отриманого в культурі жіночого диплоїдного штаму Oedogonium cardiacum більші, ніж у гаплоїдного (звичайного у природі), оогонії, в яких, як правило, виявляється більший простір між поверхнею яйця і навколишнього оогоніальной стінкою, що дає явну перевагу для спостереження початкових стадій злиття гамет в живому матеріалі. Конкретні приклади плазмогаміі і наступного етапу статевого злиття - каріогамія буде подано при описі пологів, у яких цей процес детально досліджувався.
В результаті статевого процесу утворюється зигота, яка містить одне купуляціонное диплоидное ядро - продукт злиття двох гаплоїдних ядер, що походять з тієї та іншої гамети. Зазвичай зигота оточується товстою оболонкою, переповнюється запасними продуктами і розчинною в жирах пігментом цегляно-червоного кольору - астаксантин. Зигота проростає або безпосередньо після утворення, або після більш-менш тривалого періоду спокою.
Життєвий цикл. Співвідношення диплоїдної і гаплоїдної фаз в життєвому циклі різних водоростей неоднаково. В одних випадках проростання зиготи супроводжується редукційним поділом копуляціонного ядра (зіготіческая редукція) і розвиваються при цьому рослини виявляються гаплоїдними. Так, у багатьох зелених водоростей (вольвокальние, більшість хлорококкальних, кон'югатофіціевие, харофіціевие) зигота - єдина диплоидная стадія в циклі розвитку, вся вегетативна життя проходить у них в гаплоїдному стані, вони є гаплонтамі.
В інших водоростей, навпаки, вся вегетативна життя здійснюється в диплоидном стані, а гаплоидная фаза представлена лише гаметами, перед освітою яких і відбувається редукційний розподіл ядра (гаметіческаяредукція). Зигота без редукційного поділу ядра проростає в диплоїдний таллом. Ці водорості - джлонти. Такими є багато зелені водорості, що мають сифонові будова, всі діатомові, з бурих - представники порядку фукальние.
У третіх редукційний розподіл ядра передує утворенню зооспор або апланоспор, що розвиваються, як правило, на диплоїдних таллома (споріческая редукція). Ці клітини безстатевого розмноження виростають у гаплоїдні рослини, що розмножуються тільки статевим шляхом. Після злиття гамет зигота розвивається в диплоидное рослина, що несе тільки органи безстатевого розмноження. Таким чином, у цих водоростей має місце чергування форм розвитку (генерацій): диплоїдного безстатевого спорофіта й гаплоїдного статевого гаметофіту. Обидва покоління можуть бути однакові морфологічно (ізоморфна зміна генерацій) або ж різко різні за зовнішнім виглядом (гетероморфний зміна генерацій). Ізоморфна зміна генерацій характерна для морських видів Ульв, ентероморфи, кладофори, хетоморфи із зелених водоростей, для ряду порядків бурих і більшості червоних водоростей (кілька ускладнена), гетероморфний зміна генерацій особливо поширена серед бурих водоростей, але зустрічається у зелених і червоних. Слід зазначити, що у ряду цих водоростей, що мають зміну генерацій, крім двох кардинальних моментів в їх життєвому циклі - копуляції (сіігаміі), що супроводжується подвоєнням числа хромосом в ядрі копуляціонном, і мейозу, що приводить до редукції числа хромосом, - можуть відбуватися й інші процеси : спонтанне множення числа хромосом, партеногенетичний розвиток гамет, явище гетеробластіі. У результаті спосіб розмноження (безстатеве або статевий) далеко не завжди корелює з плоїдності таллома, тобто спорофіти можуть бути не тільки диплоїдними, але і гаплоїдними і тетраплоїдні, а гаметофіти виявляють не тільки гаплоїдний, але і диплоїдний набори хромосом і т.п. Такі складні цикли розвитку були встановлені, наприклад, у ектокарпуса, ульва. Крім того, при певних зовнішніх умовах з життєвого циклу однієї і тієї ж водорості можуть випадати будь-які стадії, наприклад, у ектокарпуса відбувається реплікація спорофіта допомогою нейтральних зооспор, і в деяких місцях проживання відомі тільки диплоїдні талломи цієї водорості.
У одного й того ж виду спостерігаються різні життєві цикли. Так, у зеленій водорості Bryopsis hypnoides зустрічається як однофазний цикл, так і двофазне розвиток, у червоних водоростей Gigartina steliata, G. agardhtii відзначається як статевий цикл, так і апоміктіческое (без злиття статевих клітин) розвиток. Навіть у таких високоорганізованих форм, як ламінарія, де, як правило, регулярна зміна спорофіта й гаметофіта носить облігатний характер, можливі відхилення в життєвому циклі: партеногенетичний (з незаплідненої яйцеклітини) розвиток спорофитов і апоспоріческое (з вегетативних клітин спорофіта) розвиток гаметофитов.
Життєві цикли водоростей дуже складні і різноманітні. Особливо це відноситься до червоних водоростей, дослідження розвитку яких дозволило виявити набагато більше типів життєвих циклів, ніж це було відомо раніше. Культуральні дослідження дозволили встановити зв'язки міжводоростями, довгий час вважалися самостійними видами і опинилися лише стадіями в життєвому циклі однієї і тієї ж водорості, тобто значно розмножилися відомі нам випадки гегероморфной зміни генерацій. Це відноситься не тільки до червоних водоростей, а й до зелених - улотріхальним, бріопсідальним.
Нарешті, у небагатьох водоростей, наприклад у водорості празіоли (із зелених) - Леманн, батрахоспермума (червоні), мейоз відбувається в деяких вегетативних клітинах диплоїдного таллома (соматична редукція), з яких потім розвиваються гаплоїдні талломи. Детальніше життєві цикли будуть розбиратися при описі відповідних груп.
Особливості біології бурих водоростей.
Цікавим є відділ Бурі водорості. Серед них можна спостерігати і мікроскопічні нитчасті організми, і гіганти, що сягають 30—50 м завдовжки (наприклад, макроцистіс). Бурих водоростей існує близько 1500 видів; це фітобентос переважно холодних морів та океанів усього світу, заселяють мілководдя, але зустрічаються і далеко від берегів. У Чорному морі біля берегів України трапляються такі бурі водорості, як диктіота, цистпозира. За зовнішнім виглядом бурі водорості можуть нагадувати кущики, гіллясті шнури. Крім хлорофілу вони містять пігмент фукоксантин (бурий). Піреноїдів немає. Це найорганізованіший відділ водоростей. Талом у багатьох видів диференційований на різні тканини (асиміляційна, запаслива та механічна тканини з потовщеними стінками). Бурі водорості утворюють підводні "луки" та величезну кількість біомаси. Наприклад, це всім відома морська капуста, або ламінарія. Найскладніше організованими серед бурих водоростей є саргасуми. Ці багаторічні водорості живуть компактно в Атлантичному океані (Саргасове море). Саме тут нереститься вугор.
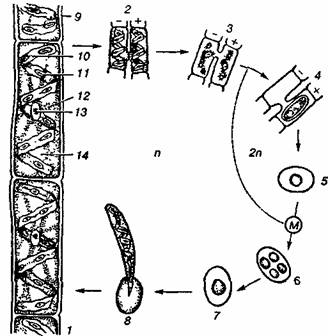
Мал. 3. Життєвий цикл спірогіри:
1 — частина талому; 2, 3 — послідовність кон'югації; 4, 5 — зиготи; 6, 7 — мейоз зиготи і відмирання трьох гаплоїдних ядер; 8 — проростання зиготи; 9 — клітинна стінка; 10 — хроматофор; 11 — піреноїд; 12— цитоплазма; 13 — ядро; 14 — вакуоля
Вегетативне розмноження відбувається частинками талому; безстатеве — зооспорами, у деяких спорами особливої будови; статеве — сперматозоїдами та яйцеклітинами.
Червоні водорості (близько 4000 видів) — багато в чому подібні до бурих. Сучасна наука виділяє їх із загальної групи водоростей в окреме підцарство, зважаючи на те, що
вони мають надто своєрідні форми спороношення і статевого розмноження. В їхньому життєвому циклі змінюється три покоління, а не два, як в інших рослин. Рухливих стадій у циклі розвитку немає. Більшість червоних водоростей — дводомні рослини. Агар-агар — це речовина, яку виробляють з багрянкового крохмалю, продукту фотосинтезу червоних водоростей. У Чорному морі зростає церамій червоний, якого хвилями викидає на берег під час шторму.
Роль водоростей у природі. Водорості — планктонні і бен-тосні, наземні й грунтові — відіграють важливу роль у природі. Разом з іншими водяними рослинами вони виробляють близько 80 % усієї маси органічних речовин, що утворюються на Землі. Серед них найпродуктивнішими є планктонні завдяки своїй здатності швидко розмножуватись.
Наземні водорості часто виступають у ролі піонерів рослинності, поселяючись на безплідних ділянках суші: скелях, пісках. У симбіозі з грибами водорості утворюють своєрідні організми — лишайники.
Водорості — одні з найдавніших організмів, що населяють нашу планету. Від них виникли наземні рослини. Збагативши атмосферу киснем, вони зумовили можливість існування різноманітного світу тварин і сприяли розвитку аеробних бактерій. Завдяки їхній діяльності в атмосфері з'явився озоновий екран, який захищає Землю від радіаційного випромінювання. Органічні речовини, які створюють водорості в процесі фотосинтезу, стають їжею для бактерій і тварин, зокрема риб.
Водорості беруть участь у колообігу речовин у природі, в поліпшенні газового режиму водойм та утворенні відкладів сапропелю (органічного мулу).
Зарості великих водоростей є укриттям для розмноження багатьох побережних тварин і дрібних водоростей. З водоростей утворились потужні поклади гірських порід: у крейдяних породах 95 % становлять рештки оболонок деяких золотистих водоростей, діатоміти на 50—80 % складаються з панцирів діатомових водоростей.
Рифи в морях і океанах також формуються за участю водоростей. Так, у рифах островів Фіджі в Тихому океані водоростей майже в 3 рази більше, ніж коралів.