ВИДЫ И РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ.
СТРОЕНИЕ СКЕЛЕТНЫХ МЫШЦ
Скелетные мышцы покрыты толстой соединительнотканной оболочкой. В толще мышцы более тонкая оболочка из соединительной ткани окружает как пучки мышечных волокон, так и отдельные волокна. Эта оболочка выполняет две функции. Во-первых, через неё к мышечным волокнам проходят нервы, кровеносные и лимфатические сосуды. Во-вторых, эластичная соединительная ткань объединяет тянущие усилия множества мышечных волокон.
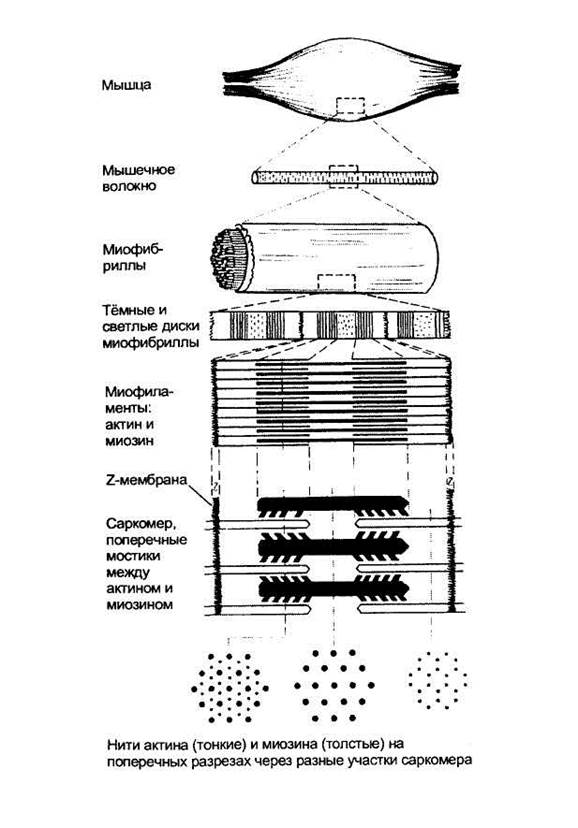
Мышечные клетки, или волокна, имеют форму заострённых на концах цилиндров. Их тонкая цитоплазматическая мембрана называется сарколеммой, а их цитоплазма - саркоплазмой. По периферии саркоплазмы располагаются многочисленные ядра мышечного волокна, а в центральной части плотно уложены ориентированные продольно миофибриллы; в одном мышечном волокне их может содержаться больше тысячи.
Именно миофибриллы создают картину поперечной исчерченности мышечного волокна, в котором чередуются тёмные и светлые диски. Тёмные диски характеризуются двойным лучепреломлением в поляризованном свете, поэтому их называют анизотропными, в отличие от светлых изотропных дисков (рис. 5). В середине каждого изотропного диска видна тёмная тонкая линия, получившая название Z-мембраны. Участок миофибриллы между соседними Z-мембранами называется саркомером, его длина в среднем составляет около 2-3 мкм.
Саркомеры образованы нитями двух сократительных белков - актина и миозина. Более тонкие нити актина одним концом прикрепляются к Z-мембране. К этой же мембране с другой стороны прикрепляются, как щетинки к щётке, актиновые нити соседнего саркомера. В промежутки между актиновыми нитями примерно на 1/4 их длины входят толстые нити второго сократительного белка - миозина. Участки миофибриллы, содержащие миозин, соответствуют тёмным анизотропным дискам, а актин - светлым изотропным дискам. Мышечное сокращение осуществляется с помощью специального механизма втягивания тонких нитей актина в центр саркомера между толстыми нитями миозина. При сокращении уменьшается только длина саркомеров, тогда как длина актиновых и миозиновых нитей не изменяется.
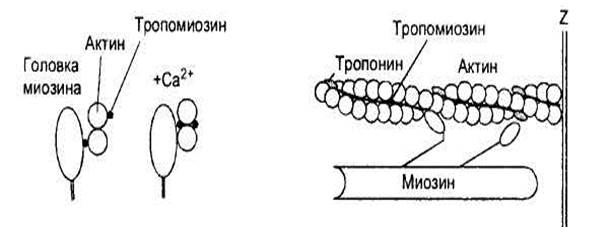
Каждая тонкая нить актина образована двойной цепью глобулярных молекул мономеров актина, что можно сравнить с двойной ниткой бус, слегка скрученных по оси (рис. 6). На каждой стороне такой двойной цепи есть продольные спиральные желобки. В желобках уложены длинные и тонкие молекулы регуляторного белка тропомиозина. К нитям тропомиозина присоединены молекулы второго регуляторного белка - тропонина, расположенные на одинаковом расстоянии друг от друга. Регуляторные белки контролируют соединения между нитями актина и миозина: от этого зависит сокращение и расслабление мышцы. Например, в расслабленной мышце молекулы тропомиозина занимают положение, препятствующее соединению миозиновых нитей с актиновыми.
|
У толстых нитей миозина есть многочисленные боковые ответвления, или поперечные мостики, имеющие форму двойных головок (рис.6). Эти головки могут поворачиваться, как на шарнирах, прикрепляться к нитям актина, а затем создавать тянущее усилие. Половина головок каждой нити миозина обращена в одну сторону, а половина - в другую, следовательно каждая половина головок может присоединиться к нитям актина, связанным как с одной, так и с другой Z-мембранами саркомера.
| В расслабленной мышце | Присоединение миозина к актину при сокращении |
Рис. 6. Сократительные и регуляторные белки миофибрилл. В расслабленной мышце тропомиозин препятствует образованию поперечных мостиков между актином и миозином. При возбуждении мышцы в саркоплазме повышается концентрация ионов кальция, которые соединяются с тропонином, что меняет конформацию его молекул. В результате этого тропонин смещает молекулу тропомио-зина в глубину желобка актиновой нити: тем самым головкам миозина предоставляется возможность соединяться с нитями актина
От сарколеммы мышечного волокна отходят внутрь узкие канальцы, получившие название поперечных Т-трубочек. Когда возбуждение передаётся через нервно-мышечный синапс на сарколемму, волна деполяризации распространяется и на Т-трубочки. В саркоплазме с Т-трубочками соседствуют цистерны саркоплазматического ретикулума, представляющего мышечный аналог гладкого эндоплазматического ретикулума. Вокруг одного саркомера проходят две Т-трубочки, а к каждой из них с обеих сторон прилежат цистерны саркоплазматического ретикулума. Каждая Т-трубочка с двумя прилежащими к ней цистернами саркоплазматического ретикулума образует триаду. Цистерны ретикулума соединены друг с другом посредством множества продольных трубочек, образуя единую систему. В саркоплазматическом ретикулуме накапливаются ионы кальция, поступающие из саркоплазмы активным транспортом. Главный интегральный белок мембраны саркоплазматического ретикулума выполняет функцию кальциевого насоса и при расслаблении мышцы перекачивает ионы кальция в ретикулум, используя для этого энергию АТФ.
НЕЙРО-МОТОРНЫЕ ЕДИНИЦЫ
Структурно-функциональной единицей скелетных мышц является многоядерное мышечное волокно. Объединяясь в пучки, эти волокна образуют мышцу. Любое мышечное волокно иннервируется только от одного мотонейрона. В то же время один мотонейрон иннервирует группу мышечных волокон, вместе с которыми он образует двигательную единицу. Она является наименьшей функциональной единицей моторной системы. В разных мышцах двигательные единицы сильно отличаются по количеству мышечных волокон. Так, например, в глазо-двигательных мышцах на один нейрон приходится около десятка мышечных волокон, в двигательных единицах мелких мышц руки их около сотни, а в крупных мышцах конечностей и туловища - более тысячи на один мотонейрон. Малые единицы характерны для мышц, с помощью которых совершаются точные, хорошо координированые движения, тогда как большие преобладают в массивных мышцах, таких например, как мышцы спины.
В зависимости от скорости развития максимальной силы сокращения, а также от быстроты утомления различают два типа двигательных единиц: 1) быстрый, утомляемый тип; 2)медленный, мало утомляемый. В большинстве мышц представлены два типа двигательных единиц, но в разном соотношении, которое варьирует у людей индивидуально. В спортивной физиологии людей с относительно большим процентом быстрых волокон относят к спринтерам, а обладателей большего, чем в среднем, процента медленных волокон - к стайерам. У всех людей при старении процент быстрых волокон уменьшается.
Волокна быстрых двигательных единиц относительно меньше снабжаются кровью и поэтому имеют бледно-красный цвет. Они предназначены для мощных быстрых сокращений в течение короткого времени, поскольку быстро утомляются, не поспевая из-за относительно малого кровоснабжения окислять глюкозу до конечных продуктов. Волокна медленного типа обеспечены обильной сетью кровеносных капилляров, что придаёт им тёмно-красный цвет. И, хотя они сокращаются медленнее, зато способны очень долго работать без утомления. Развиваемая ими сила составляет от 1 до 10% от силы быстрых волокон.
Тела мотонейронов двигательных единиц медленного типа имеют наименьшие размеры и самый низкий порог возбуждения, поэтому только они могут активироваться слабыми входными сигналами. Мотонейроны быстрых двигательных единиц, напротив, являются самыми крупными и, чтобы возбудить их, нужны сильные стимулы. Поэтому медленные двигательные единицы используются при любых движениях, а быстрые - только в тех случаях, когда понадобится большая сила сокращающихся мышц. Так, например, при стоянии или спокойной ходьбе сокращение мышц обеспечивает менее чем половина всех двигательных единиц - здесь сила мышечного сокращения составляет около 20% от максимальной. Для того чтобы выполнить прыжок, может потребоваться максимальная сила: в таком случае понадобится активация мотонейронов быстрых двигательных единиц.
ВИДЫ И РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ.
Мышечное сокращение проявляется в укорочении мышцы. В зависимости от условий стимуляции и функционального состояния мышцы может возникнуть одиночное и тетаническое сокращение мышцы.
Амплитуда одиночного сокращения зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных волокон, составляющих целую мышцу, различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна.
Мышечному сокращению предшествует процесс возбуждения (т.е. ПД), который совпадает по времени с латентным периодом мышечного сокращения.

Рис. 7. Соотношения фаз сократимости(А) и возбудимости(Б) поперечно-полосатых мышц:
1,6 - исходная возбудимость; 2,5 - супернормальная возбудимость; 3 - абсолютная рефрактерность; 4 - относительная рефрактерность
Сила и скорость мышечного сокращения пропорциональны также частоте потенциалов действия, распространяющихся к мышце по аксону мотонейрона - это ещё один механизм регуляции, который называется частотным кодированием. Потенциал действия в нерве или мышце длится приблизительно 1-3 мс, а самое короткое сокращение, состоящее из укорочения и расслабления, продолжается примерно 100 мс (рис 7). Таким сокращением мышца отвечает на единичный потенциал действия - это сокращение называется одиночным (рис.8).

Рис.8 - Одиночное мышечное сокращение.
1 - латентная фаза; 2 - фаза сокращения; 3 - фаза расслабления
Если частота возбуждающих мышцу нервных импульсов станет расти, она не успеет полностью расслабиться к моменту прихода очередного потенциала действия, и сила её сокращения будет увеличиваться. Такой вид сокращения получил название зубчатого тетануса. С ещё большим увеличением частоты стимуляции формируется плато: в это время мышца совсем не расслабляется, а сокращение делается максимальным - этот режим называется гладким тетанусом (рис.9).
|

А. При малой частоте нервных импульсов, возбуждающих мышцу, она отвечает на каждый из них одиночным сокращением, успевая расслабиться до прихода следующего импульса;
Б. С увеличением частоты нервных импульсов происходит суммация одиночных сокращений, при которой растёт амплитуда сократительных ответов;
В. При достаточно большой частоте нервных импульсов мышца не может расслабиться, а сократительный ответ становится максимальным.
Серия следующих друг за другом потенциалов действия приводит к значительному повышению внутриклеточной концентрации кальция, поэтому сократительный ответ и становится сильнее и продолжительнее. Частота поступающих к мышцам потенциалов действия варьирует в небольших пределах. Например, при произвольных сокращениях потенциалы действия в мотонейронах появляются с частотой приблизительно 8 Гц, в редких случаях она может превысить 25 Гц. С нарастанием частоты поступающих к мышце потенциалов действия сила её сокращения увеличивается (рис.10).
|
