Природа и состав ферментов. Апофермент, кофактор. Коферменты и простетические группы. Функции белковой и небелковой части сложных ферментов.
Доказательства белковой природы ферментов:
1. Ферменты при гидролизе распадаются на аминокислоты.
2. Под действием разл. факторов ферменты подвергаются денатурации и теряют активность.
3. Осуществлено выделение ферментов в форме кристаллов белка.
4. Ферменты оказывают высокоспецифическое действие.
Большинство ферментов имеют четвертичную структуру: функционирование такого белка нуждается не в одной, а в нескольких молекулах (чаще всего в двух или четырёх), которые вместе образуют комплекс, обладающий всеми специфическими свойствами. Каждая отдельная молекула такого белка называется субъединицей. В одних случаях субъединицы сами обладают активностью, в других - субъединицы по отдельности неактивны.
Состав ферментов.По химической природе фермент могут быть белками простыми (ферменты протеины) и сложными (ферменты протеиды - содержат в своём составе, помимо белкового компонента, ещё и небелковый, например, нуклеотиды, витамины, атомы (катионы) металла).
Большинство природных ферментов - сложные белки – протеиды. Протеиды состоят из:
Апофермент (белковая часть) + кофактор (небелковая часть) = холофермент (активный комплекс).
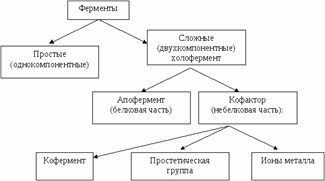 Кофактор (небелковые компоненты - коферменты и простетические группы и ион металлов) чаще всего представлен витаминами или соединениями, построенными с их участием (коэнзим А, НАД+, ФАД); фосфорные эфиры некоторых моносахаридов, или ионами металлов, и необходим для выполнения ферментом его каталитической роли. Кофакторы могут иметь различную химическую природу и различаться по прочности связи с полипептидной цепью .
Кофактор (небелковые компоненты - коферменты и простетические группы и ион металлов) чаще всего представлен витаминами или соединениями, построенными с их участием (коэнзим А, НАД+, ФАД); фосфорные эфиры некоторых моносахаридов, или ионами металлов, и необходим для выполнения ферментом его каталитической роли. Кофакторы могут иметь различную химическую природу и различаться по прочности связи с полипептидной цепью .
Классификация кофакторов
Все кофакторы (коферменты и простетические группы) – это низкомолекулярные органические соединения. В настоящее время принята классификация кофакторов по функциональному признаку. В соответствии с этим все кофакторы делят на три группы:
1. кофакторы окислительно-восстановительных процессов: (NAD,NADР), флавинмононуклеотид (FMN), флавинадениндинуклеотид (FAD), железопорфирины, убихинон, аскорбиновая кислота;
2. кофакторы переноса групп: нуклеозидфосфаты, фосфаты сахаров, коэнзим А (СоА, HSCoA, фолиевая кислота, пиридоксальфосфат.;
3. кофакторы процессов синтеза, изомеризации и расщепления С-С связей: тиаминдифосфат, биотин, глутатион, кобамидные коферменты.
Кофермент - небелковый фактор, который легко отделяется от белковой части - апоферментапри диссоциации.
Простетическая группа– ковалентно связанный с белковой цепью небелковый компонент, который не отделяется при выделении и очистке фермента.
Кроме этого, роль кофактора могут выполнять металлы: Mg2+, Мn2+, Са2+и др.
Важнейшие коферменты и простетические группы ферментов
NAD - Никотинамид (витамин РР), FAD - Рибофлавин (витамин В2), Коэнзим А (СоА) - Пантотеновая кислота
Отличительная особенность двухкомпонентных ферментов: ни кофактор отдельно (включая большинство коферментов), ни сам по себе апофермент каталитической активностью не наделены, и только их объединение, протекающее не хаотично, а в соответствии с программой их структурной организации, обеспечивает быстрый ход химической реакции.
Функции коферментов и простетической группы следующие:
· участие в акте катализа,
· осуществление контакта между ферментом и субстратом,
· стабилизация молекулы фермента в пространстве.
Коферменты можно рассматривать как составную часть молекулы фермента.
Функции апофермента:
· усиливает каталитический акт небелковой части,
· определяет специфичность ферментов, поскольку одна и та же по химизму небелковая часть может функционировать в составе различных ферментов. Так, никотинамид (НАД +) является коферментом многих дегидрогеназ.
· обеспечивает специфичное опознавание субстратов, приводящее к образованию комплекса
· обеспечивает связывание в активном центре иона или молекулы кофактора.
Большинство коферментов не синтезируются в организме млекопитающих. Они должны поступать в организм с пищей (как правило, растительной). Однако в организм попадают не сами коферменты, а их предшественники — витамины. В клетке витамины модифицируются до коферментной формы.
Ионы металла в составе ферментов:
Многие ферменты (около 2/з) являются металлоферментами:
для активации ферментов свертывания крови требуется Са2+;
оксидоредуктазы используют в качестве кофакторов Fe2+, Cu2+, Mn2+;
киназы — Mg2+;
для глутатионпероксидазы (важного фермента в системе обезвреживания активных форм кислорода) требуется Se.
Этим объясняется, что в любой диете должны присутствовать эти и другие микроэлементы.
Понятным становится токсический эффект тяжелых металлов, например Cd2+, Hg2+, которые могут замещать Zn2+ в активном центре определенных ферментов, включая РНК-полимеразу, уменьшая активность ферментов.
Механизм действия ферментов. Активный центр фермента и его строение. Роль фермент-субстратного комплекса (общие представления)
В процессе катализа реакции в контакт с субстратом вступает не вся молекула фермента, а определенный ее участок, который называется активным центром - это участок в пространственной структуре фермента, с которым связывается субстрат и подвергается химическому превращению. Эта зона молекулы не состоит из последовательности аминокислот, а формируется при скручивании белковой молекулы в третичную структуру. Отдельные участки аминокислот сближаются между собой, образуя определенную конфигурацию активного центра. Важная особенность строения активного центра - его поверхность комплементарна поверхности субстрата, т.е. остатки АК этой зоны фермента способны вступать в химическое взаимодействие с определенными группами субстрата. Число активных центров может быть равно числу субъединиц в четвертичной структуре фермента, т.е. сколько субъединиц, столько активных центров.
Особенности образования активного центра у простых белковых ферментов.
Обычно он образован 12-16 аминокислотными остатками полипептидной цепи. Иногда их число больше. Аминокислоты, формирующие активный центр, находятся в разных местах полипептидной цепи. При пространственной укладке белка-фермента (в третичную структуру), они сближаются и образуют активный центр.
Особенности образования активного центра у ферментов-протеидов (сложных белков-ферментов).
 У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора.
У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора.
В активном центре условно выделяют два участка:
- якорный (контактный, связывающий), отвечающий за специфичность связывания субстрата (узнавание);
- каталитический, где происходит химическое превращение субстрата после его связывания (сначала фермент узнает субстрат, притягивает его, затем субстрат располагается в этом активном центре).
Аллостерический центр (allos– чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Аллостерические ферменты в большинстве случаев являются белками с четвертичной структурой, при этом активный и регуляторный центры могут находиться в разных субъединицах.
У сложных ферментов в формировании активных центров принимают участие также функциональные группы коферментов.
Механизм действия простого и сложного ферментов одинаков, так как активные центры в их молекулах выполняют сходные функции.
В основе действия ферментов лежит их способность ускорять реакции за счет уменьшения энергии активации субстрата. Ферменты деформируют электронные оболочки субстратов, облегчая таким образом взаимодействие между ними. Энергия, необходимая для того, чтобы привести молекулы в активное состояние, называется энергией активации. Роль обычного катализатора (и еще в большей мере биологического) состоит в том, что он снижает энергию активации субстрата.
 Основы механизма действия ферментов были изучены в начале XX в.
Основы механизма действия ферментов были изучены в начале XX в.
Существует две модели, описывающие взаимодействие субстрата с активным центром:
 а) Модель жёсткого соответствия («ключ – замок»), предложена Э. Фишером в 1890 году. Активный центр считается заранее подогнанным под форму молекулы субстрата. Эта модель не утратила своего значения для понимания некоторых свойств ферментов, например, их способности к строго определённому связыванию двух или большего числа субстратов или для объяснения кинетики насыщения субстратом.
а) Модель жёсткого соответствия («ключ – замок»), предложена Э. Фишером в 1890 году. Активный центр считается заранее подогнанным под форму молекулы субстрата. Эта модель не утратила своего значения для понимания некоторых свойств ферментов, например, их способности к строго определённому связыванию двух или большего числа субстратов или для объяснения кинетики насыщения субстратом.
б) Модель индуцированнного соответствия («рука – перчатка»), предложена Кошлендом в 1950-е годы. Согласно этой модели, субстрат вызывает (индуцирует) конформационные изменения фермента, и лишь в результате этих изменений аминокислотные остатки фермента принимают пространственную ориентацию, необходимую для связывания субстрата и катализа. При этом другие аминокислотные остатки могут погрузиться вглубь молекулы фермента