Механорецепторы дыхательной системы
Механорецепторы дыхательной системы выполняют двоякую роль: во—первых, они участвуют в регуляции параметров дыхательного цикла — глубины вдоха и его длительности; во—вторых, эти рецепторы служат источником ряда рефлексов защитного характера — кашля, например.
Рецепторы растяжения легких. Эти рецепторы локализованы главным образом в гладкомышечном слое стенок трахеобронхиального дерева и чувствительны к трансмуральному давлению, т. е. к разности давлений внутри и снаружи просвета воздухоносных путей. Тем самым эти Механорецепторы оказываются источником сигнализации о растяжении дыхательных путей, а следовательно, и легких. Афферентные волокна рассматриваемых рецепторов идут в составе блуждающего нерва, центральный путь их импульсации прослежен до области дорсальной дыхательной группы ядер продолговатого мозга.
Возбуждение рецепторов растяжения легких, нарастая в ходе вдоха вызывает в конечном счете торможение активности инспираторных нейронов центрального дыхательного механизма, способствуя прерыванию вдоха и смене его выдохом (рефлексы Геринга—Брейера). Так как активность одной части этих рецепторов («статических») зависит от достигнутого легочного объема а другой части («динамических») — от скорости вдоха, то прекращение вдоха наступает тем скорее, чем глубже данный вдох и чем быстрее он развивается. Так регулируется паттерн дыхания — соотношение между глубиной и частотой дыхания.
Роль рецепторов растяжения легких наглядно выявляется при их выключении с помощью блокады или перерезки блуждающих нервов (ваготомии): вдохи становятся углубленными, затянутыми, как это происходит при разрушении пневмотаксического центра. Если же разрушение названного центра сочетать с ваготомией, наступает апнейзис: дыхательные движения останавливаются на вдохе, который лишь изредка прерывается короткими экспирациями.
Ирритантные рецепторы. Ирритантные (от лат. irritatio — раздражать) рецепторы расположены в эпителиальном и субэпителиальном слоях стенок воздухоносных путей. Их афферентные волокна тоже идут в стволе блуждающего нерва. Эти рецепторы реагируют на резкие изменения объема легких, в частности на их спадение, которое вызывает рост инспираторной активности центрального механизма, прерывая таким путем выдох. Чувствительны они и к частицам пыли, скоплению слизи некоторым химическим раздражителям. По—видимому, возбуждение ирритантных рецепторов характерно для многих патологических изменений в легких и воздухоносных путях.
Юкстаальвеолярные (юкстакапиллярные), или J—рецепторы. Эти рецепторы чувствительны к ряду биологически активных веществ (никотину, гистамину, простагландинам и др.), проникающим либо из воздухоносных путей, либо с кровью малого круга. Они локализованы в интерстиции легких вблизи капилляров альвеол и дают начало немиелинизированным С—волокнам, проходящим в составе блуждающего нерва. Стимуляция ирритантных и J—рецепторов проявляется в частом, поверхностном дыхании (тахипноэ), которое наблюдается, например, при ограничении (рестрикции) газообменной поверхности легких.
Рецепторы верхних дыхательных путей. Они служат главным образом источником рефлексов защитного характера, возникающих при скоплении в воздухоносных путях слизи, попадании инородных тел и химических раздражителей. Эти реакции опосредованы афферентными волокнами целого ряда черепных нервов: верхнего гортанного (ветвь блуждающего нерва), языкоглоточного, тройничного и др. (проявляются в виде кашля, чихания, глотания и др.)
Проприоцепторы дыхательных мышц. Эти рецепторы контролируют соответствие сокращений «заданию», поступающему по эфферентным путям от центрального дыхательного механизма. В этом отношении важную роль выполняют мышечные веретена межреберной и брюшной мускулатуры. Если укорочение мышцы оказывается меньше заданного (например, вследствие повышенного сопротивления дыханию), импульсация от интрафузальных волокон усиливается и через посредство —петли повышается активность соответствующих спинальных —мотонейронов. Сокращение мышцы усиливается, что способствует преодолению возникшего препятствия. Правда, основной инспиратор — диафрагма — бедна мышечными веретенами, здесь преобладают сухожильные рецепторы Гольджи. Функцию афферентной системы, контролирующей ее сокращения по принципу обратной связи, выполняют описанные выше механорецепторы легких и воздухоносных путей.
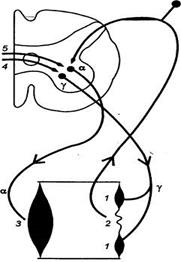
|
| Рис. Связи мышечных веретен с сегментарными мотонейронами спинного мозга Рассогласование между укорочением интрафузальных (1) и экстрафузальных (3) волокон мышцы, иннервируемых соответственно альфа () и гамма () —мотонейронами, вызывает стимуляцию аннулоспирального окончания (2), что ведет к дополнительной активации альфа—мотонейрона. Таким способом, в частности, происходит компенсация увеличенного сопротивления дыханию путем усиления сокращения межреберных мышц и их укорочения до величины, заданной дыхательным центром через посредство прямых (5) и непрямых (4} эфферентных путей. |
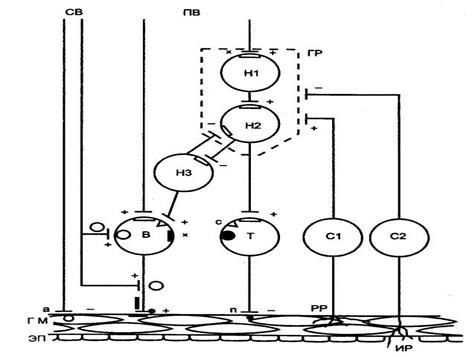
|
| Рис. Схема функционального модуля респираторной части метасимпатической нервной системы СВ и ПВ — симпатические и парасимпатические нервные волокна; ГР — генератор ритма; HI, H2 — нейроны генератора ритма; НЗ — интернейрон; В — возбуждающий эффекторный нейрон; Т — тормозный эффекторный нейрон; Cl, C2 — сенсорные клетки; РР — рецепторы растяжения; ИР — ирритантные рецепторы; ГМ — гладкомышечные клетки; ЭП — эпителий; «+» — возбуждающая передача; «—» — тормозная передача; х — холинергические, а — адренергические, п — пептидергические, с — серотонергические синапсы. |
Нейроны респираторной части метасимпатической нервной системы Согласно представлениям, выдвинутым в начале XX столетия Дж. Ленгли клетки интрамуральных ганглиев рассматривались как периферические (постганглионарные) нейроны парасимпатической нервной системы, через которые осуществляется холинергическая иннервация гладкой мышцы. По существующим современным взглядам, основанным на прямых электрофизиологических показателях функции этих клеток, совокупности нейронов интрамуральных ганглиев представляют собой низшие рефлекторные центры, способные к самостоятельной интегративной деятельности.
Функциональная структура этих центров представляется следующим образом. Составляющие центр нейроны объединены в функциональные модули, включающие в себя четыре основные группы клеток: формирующие ритмическую активность (генератор ритма), эффекторные возбуждающие и тормозящие мышцу и другие функции нижних дыхательных путей, а также сенсорные. В состав каждой из этих групп может входить несколько клеток. Совокупность таких функциональных модулей, объединенных возбуждающими и тормозными связями, в целом образует периферический нервный центр, управляющий, например, гладкомышечной стенкой всех нижних дыхательных путей. В сфере управления функционального метасимпатического модуля находится не только механика гладкой мышцы, но и функции секретирующего и всасывающего эпителия, местного капиллярного кровотока, местных эндокринных элементов и иммунных структур.
ТРАНСПОРТ ГАЗОВ КРОВЬЮ.
Кровь доставляет тканям кислород и уносит углекислый газ.
Движение газов из окружающей среды в жидкость и из жидкости в окружающую среду осуществляется благодаря разности их парциального давления. Газ всегда диффундирует из среды, где имеется высокое давление, в среду с меньшим давлением.
Парциальное давление кислорода в атмосферном воздухе 21,1 кПа (158 мм рт. ст.), в альвеолярном воздухе — 14,4—14,7 кПа (108—110 мм рт. ст.) и в венозной крови, притекающей к легким,—5,33 кПа (40 мм рт. ст.). В артериальной крови капилляров большого круга кровообращения напряжение кислорода составляет 13,6—13,9 кПа (102—104 мм рт. ст.), в межтканевой жидкости — 5,33 кПа (40 мм рт. ст.), в тканях — 2,67 кПа (20 мм рт. ст.). Таким образом, на всех этапах движения кислорода имеется разность его парциального давления, что способствует диффузии газа.
Движение углекислого газа происходит в противоположном направлении. Напряжение углекислого газа в тканях — 8,0 кПа и более (60 и более мм рт. ст.), в венозной крови — 6,13 кПа (46 мм рт. ст.), в альвеолярном воздухе — 0,04 кПа (0,3 мм рт. ст.). Следовательно, разность напряжения углекислого газа по пути его следования является причиной диффузии газа от тканей в окружающую среду.
Транспорт кислорода кровью. Кислород в крови находится в двух состояниях: физическом растворении и в химической связи с гемоглобином. Гемоглобин образует с кислородом очень непрочное, легко диссоциирующее соединение - оксигемоглобин: 1г гемоглобина связывает 1,34 мл кислорода. Максимальное количество кислорода, которое может быть связано 100 мл крови, -кислородная емкость крови(18,76 мл или 19 об%).
Насыщение гемоглобина кислородом колеблется от 96 до 98%. Степень насыщения гемоглобина кислородом и диссоциация оксигемоглобина (образование восстановленного гемоглобина) не находятся в прямой пропорциональной зависимости от напряжения кислорода. Эти два процесса не являются линейными, а совершаются по кривой, которая получила название кривой связывания или диссоциации оксигемоглобина.

Кривые диссоциации оксигемоглобина в водном растворе (I) и в крови (II) при напряжении углекислого газа 5,33 кПа (40 мм рт. ст.) (по Баркрофту).
При нулевом напряжении кислорода оксигемоглобина в крови нет. При низких значениях парциального давления кислорода скорость образования оксигемоглобина невелика. Максимальное количество гемоглобина (45— 80%) связывается с кислородом при его напряжении 3,47—6,13 кПа (26—46 мм рт. ст.). Дальнейшее повышение напряжения кислорода приводит к снижению скорости образования оксигемоглобина .
Сродство гемоглобина к кислороду значительно понижается при сдвиге реакции крови в кислую сторону, что наблюдается в тканях и клетках организма вследствие образования углекислого газа
Переход гемоглобина в оксигемоглобин и из него в восстановленный зависит и от температуры. При одном и том же парциальном давлении кислорода в окружающей среде при температуре 37—38° С в восстановленную форму переходит наибольшее количество оксигемоглобина,
Транспорт углекислого газа кровью. Углекислый газ переносится к легким в форме бикарбонатов и в состоянии химической связи с гемоглобином (карбогемоглобин).
.