Будова плазматичної мембрани
Відповідно до сучасних досліджень мембрана має рідинно-мозаїчну структуру (С. Сінгер і Дж. Нікольсон, 1972) (рис. 2).
В основі мембранної матриці лежить двошарова ліпідна структура. Більшу частку мембранних ліпідів складають фосфоліпіди, які здатні утворювати бішар, що складається з внутрішньої гідрофобної області (аліфатична частина жирних кислот або стероїдного кістяка холестерину) і гідрофільних поверхонь (гліцерин, залишок фосфорної кислоти, аміноспирту, OH-жирної кислоти). Тому через мембрани можуть проникати жиророзчинні речовини, а водорозчинні речовини і гідрофільні іони не в змозі перебороти гідрофобну область; вони попадають усередину клітини по спеціальних каналах проникності білкової природи. Основну частину ліпідів, що містяться в біомембранах, являють собою полярні ліпіди (гліцерофосфоліпіди, сфінгофосфоліпіди, гліцерогліко-ліпіди, сфінгогліколіпіди). Окремі фрагменти молекул фосфоліпідів чинять тепловий рух, що відіграє важливу роль: 1) у взаємодії ліпідів з білками; 2) у процесах транспорту речовин через мембрану; 3) у проникності (рис. 2).
Підвищенню «плинності» мембрани сприяє наявність у молекулі ліпідів коротколанцюжкових жирних кислот і розгалужених ланцюгів.
Чим вищий ступінь їхньої ненасиченості, тим сильніше виражений ступінь розрідженості мембрани. Ферментативна активність і пасивний транспорт тісно пов'язані з плинністю мембранних ліпідів.
Білки, в основному, представлені глікопротеїдами, розділили на два види: периферичні й інтегральні. Периферичні білки легко екстрагуються з плазматичної мембрани водяними, що не містять поверхнево активних речовин (ПАР), розчинниками. Інтегральні—тісно пов'язані з мембраною і легко можуть бути виділені за допомогою ПАР, що руйнують ліпідний бішар.
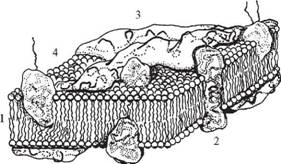
Рис- 2- Узагальнена модель типової плазматичної мембрани:
1 — ліпіди; 2 — інтегральні білки; 3 — периферичні білки; 4 — глікопротеїди
Так як полярні ліпіди, периферичні білки також мають одну полярну область і одну неполярну (домен). У полярній області білкової глобули, що знаходиться в контакті з водою, зібрані іонізовані залишки амінокислот і всі ковалентно зв'язані вуглеводні залишки. У неполярній області відсутні іонізовані і вуглеводні залишки. Ця область білкової глобули занурена в гідрофобну внутрішню частину мембрани. Інтегральні білки, що пронизують мембрану, закріплені так, що їх полярні ділянки повернені всередину і назовні, а область гідрофобного центру розташовується між ними. Інтегральні мембранні білки при фізіологічній температурі дифундують за рахунок звичайного теплового руху уздовж шару. Насичені аліфатичні ланцюги і стерини викликають підвищення в'язкості мембрани й обмежують латеральний рух молекул білка в порожнині мембрани. Мембранні білки можуть також бути обмежені в рухливості в зв'язку з присутністю пов'язаних із внутрішньою поверхнею мембрани цитоскелетних структур.
Класифікація мембранних білків у залежності від їхніх функцій
1. Ферменти—каталізатори.
2. Транспортні білки: селективні фільтри (канали в біомембранах,
вибірних стосовно іонів, що структурно відповідні до найвужчої частини каналу—фільтра); «ворота» (у збуджених мембранах при зниженні потенціалу спокою нижче порогової величини відкриваються канали й іони №+ надходять до клітини, у стані спокою в нервовому волокні №+-канали закриті «воротами»); насоси — транспортні АТФази (здійснюють перенесення речовин проти концентраційного Градієнта з витратою енергії).
3. Рецептори — це білки, що звичайно складаються з декількох доменів. Вони містять сполучнуділянку, специфічнудля природного ліганду (гормону, медіатора антитіла й ін.), тобто «пізнають» цей ліганд і взаємодіють з ефекторними системами, для яких вони також мають ділянку «пізнавання». Інформація для активації ефекторної системи цілком міститься в рецепторі мембрани. За утворенням комплексу рецептор — ліганд йдуть специфічні реакції такі, як регуляція ферментативної активності за допомогою вивільнення вторинних хімічних посередників (цАМФ, цГМФ)—циклічних нуклеотидів, відкривання або закривання іонних каналів, метилирування, фосфоритування і збільшення син-тезуДНК.
4. Поверхневі антигени (антигенні детермінанти) являють собою глікопротеїд-ліпідні комплекси (наприклад, групові антигени АВ0, Ші та інших групових систем крові, локалізовані в мембрані еритроцитів; тканинні антигени), що забезпечують імунохімічні функції.
5. Імуноглобуліни (так, у клітинній мембрані В-лімфоцитів убудо-вані молекули імуноглобулінів, які служать рецептором для специфічних антигенів). Основна функція таких білків — участь в імунологічних реакціях, тобто в створенні захисних сил організму шляхом утворення антитіл.
6. Структурні білки (спектрин у мембрані еритроцитів) відіграють важливу роль у підтриманні разом з білками цитоскелета форми клітин.
7. Скорочувальні білки, що мають АТФазну активність, тобто здатність розщеплювати АТФ із утворенням АДФ і фосфату, беруть участь у процесі екзоцитозу.
Функції плазматичної мембрани
Мембрана є не лише бар'єром між клітиною і зовнішнім середовищем, але являє собою пристрій, що забезпечує відносну сталість складу внутрішньоклітинного об'єму. Поряд з цим вона містить специфічні рецептори для зовнішніх «сигналів», присутністю яких можуть пояснюватися такі різноманітні відповіді, як орієнтований рух клітини (хемотаксис), стимуляція пов'язаних із мембраною ферментів (циклаз) чи генерації сигналів, що можуть бути хімічними (цАМФ, цГМФ) чи елек
тричними, як у нервовій клітині. Плазматична мембрана також є місцем розташування специфічних для клітини антигенів, що характерні як для даного типу клітин, так і для виду ссавця в цілому. Білки і специфічні ферменти, локалізовані в мембрані, пов'язані з транспортом іонів і метаболітів через мембранний бар'єр. Завдяки інвагінаціям (впинанням) плазматична мембрана може утворювати єдине ціле з довгими внутрішньоклітинними каналами, що дозволяє здійснювати надходження речовин ззовні далеко «всередину» клітини, і бере участь у формуванні простору між внутрішньою і зовнішньою мембранами ядра.