Нейрохимические механизмы памяти. “Ранние” и “поздние” гены. “Перенос” памяти
Постоянное хранение информации связано с химическими или структурнымиизменениями в мозгу. Практически все согласны с тем, что запоминание осуществляется посредством электрической активности, т.е. химические или структурные изменения в мозге должны каким-то образом влиять на электрическую активность.
Если системы памяти являются результатом электрической активности, то, следовательно, мы имеем дело с нервными цепями, способными реализовать следы памяти. Простейшая цепь, обеспечивающая память, представляет собой замкнутую петлю. Возбуждение последовательно обходит весь круг и начинает новый. Такой процесс называется реверберацией. Поступающий сенсорный сигнал вызывает последовательность электрических импульсов, которая сохраняется неопределенно долгое время после того, как сигнал прекратится.
Существует несколько гипотез о том, что приводит к прекращению реверберации:
Во-первых, группы клеток реверберирующей цепи в действительности расположены значительно более сложным образом. Фоновая активность этих нейронов, а также воздействия со стороны многочисленных, внешних по отношению к данной петле входов, в конечном итоге, нарушают характер циркуляции импульсов.
Во-вторых, еще один возможный механизм прекращения реверберации – появление новых сигналов, которые могут активно затормозить предшествующую реверберирующую активность.
В-третьих, импульс, поступающий в одно звено цепи, не всегда может оказаться способным вызвать активность в следующем звене и, в конце концов, поток импульсов угасает.
В-четвертых, реверберация может прекратиться вследствие какого-либо «химического» утомления в нейронах и синапсах.
Избирательная электрическая активация определенной нервной петли обеспечивает кратковременное запоминание. Как же представить в подобной схеме долговременную память? Согласно одной из популярных теорий, многократная электрическая активность в нейронных цепях вызывает химические или структурные изменения в самих нейронах, что приводит к возникновению новых нейронных цепей. Это изменение цепи называется консолидацией. Консолидация следа происходит в продолжение длительного времени. В основе долговременной памяти лежит постоянство структуры нейронных цепей.
Таким образом, кратковременная и долговременная память могут быть связаны с одними и теми же нервными элементами, с той разницей, что кратковременная память – это временная электрическая активность определенных нейронов, а долговременная память – постоянная структура тех же самых нейронов.
Существуют 2 гипотезыотносительно механизмов консолидации:
Первая предполагает, что долговременная память заключена в структуре белковых молекул в каждом синапсе. И нервная информация переходит через синаптическую щель химическим путем.
Согласно другой точке зрения долговременная память может быть результатом возникновения новых синапсов. Это означает, что всякий раз при заучивании нового материала в мозгу возникают физические изменения. Но микроскопической техникой эти изменения обнаружить не удается, в частности вследствие исключительной трудности наблюдения живых нервных клеток под микроскопом.
После того, как были открыты химические процессы, лежащие в основе наследственности, возникла мысль, что те же самые механизмы могли бы участвовать в процессах запоминания. Поскольку ДНК содержит генетическую память для каждого индивидуального организма, логично предположить, что она или РНК может также передавать и приобретенный опыт.
Инструкции для синтеза белка, переносимые молекулой РНК, заключены в специфической последовательности органических оснований, присоединенных к остову молекулы, именно они служат матрицами для синтеза белков. Различная последовательность приводит к синтезу разных белков. Можно предположить, что эта последовательность изменяется в результате опыта, приобретенного животными при обучении. + см.вопрос 37
Молекулярные механизмы долговременной памяти (экспрессия генов)
Эффекторные белки определяют хранение информации в организме,
Регуляторные белки – присоединяясь к ДНК или отделяясь от нее, контролируют экспрессию генов.
Внешнее воздействие приводит к изменению внеклеточной среды, вызывает в геноме каскадную реакцию, в которой выделяют две фазы:
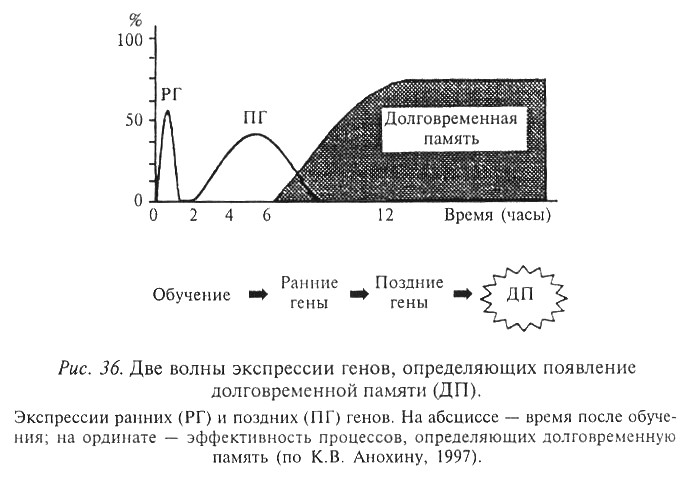
1. Фаза активации: соответствует индукции специфических регуляторных генов из класса ранних генов.Известно около 100 ранних генов. Продуктами большинства из них являются регуляторные белки.
Активность этих белков наступает через 15-30 мин после воздействия, кратковременна (от 1 до 3-х часов) – этот процесс соответствует кратковременной и промежуточной памяти (консолидации).
Ранние гены контролируют транскрипцию поздних генов, которые являются для них мишенями. Регуляторные белки (продукты ранних генов) продуцируют экспрессию поздних генов – морфорегуляторных. Эти гены определяют вторую фазу синтеза РНК и белков.
2. Вторая фаза синтеза РНК и белков – обуславливает рост и изменение клеточных связей в структурах мозга. Вторая волна активности появляется через 3 часа после воздействия и длится около 5 часов.
Включает синтез 4-х новых белков, через 24 часа – еще 2-х белков.
Долговременная памятьформируется после экспрессии поздних генови зависит от индукции новых генов через вторичных посредников.
Предполагают, что ранние эффекторные гены отвечают за синтез белков, которые сохраняют информацию в течение дней.
Поздние эффекторные гены поддерживают информацию в течение недель и месяцев.
Более подробное объяснение этой схемы:
Формирование нового опыта требует экспрессии генов в мозге.
Основным шагом в понимании биологических механизмов консолидации памяти стало открытие 1960-х годов, показавшее, что переход памяти из кратковременной в долговременную форму требует синтеза новых молекул РНК и белка, т.е. экспрессии генов. Было установлено, что волна синтеза новых белков в клетках при запоминании информации совпадает с периодом консолидации памяти, а химическая блокада экспрессии генов в этот период нарушает образование долговременной памяти.
Оказалось также, что "критическое окно" амнестического действия блокаторов экспрессии генов универсально для самых разных видов обучения и различных организмов, от беспозвоночных до человека. Данное предположение также хорошо согласовывалось с гипотезой об участии клеточного роста и изменения морфологии синапсов в долговременной памяти.
Таким образом, понятие долговременной памяти постепенно трансформировалось из условного обозначения относительной продолжительности явления, в компонент биологической концепции, связывающей научение и опыт с морфогенезом и развитием.
Критическим звеном этой концепции стал молекулярный механизм консолидации памяти, отождествляемый с активацией транскрипции генов в нервных клетках при научении. Однако то, какие именно гены активируются при научении и каковы их функции в нервных клетках, долгое время оставалось неизвестным.
При научении в мозге активируются гены транскрипционных факторов. Первыми генами, активация которых была обнаружена в мозге при обучении, оказались так называемые "непосредственные ранние гены", кодирующие транскрипционные факторы. "Непосредственные ранние гены" (НРГ) были впервые обнаружены при изучении механизмов геномного ответа на действие факторов роста, запускающих процессы клеточного цикла. Индукция их транскрипции происходила, несмотря на подведение ингибиторов синтеза белка, то есть строилась на механизмах, заранее готовых для восприятия экстраклеточных стимулов. Первые из идентифицированных продуктов генов данного семейства оказались ядерными белками, связывающимися с ДНК и регулирующими транскрипцию других генов.
По этим свойствам данные гены значительно напоминали группу "непосредственных ранних генов" бактериофагов и эукариотических ДНК-вирусов, поэтому, по аналогии с вирусными генами, эта группа быстро активирующихся генов получила название "клеточных непосредственных ранних генов". Это же семейство часто обозначается как "гены первичного ответа", "гены раннего ответа" или просто "ранние" гены.
Одним из первых в данной группе был клонирован ген c-fos. Его структура и свойства хорошо изучены, и он может служить прототипом генов данного семейства. Первоначально было установлено, что в ходе эмбрионального развития c-fos играет важную роль в регуляции процессов клеточного роста и пролиферации. Гены, экспрессия которых находится под контролем индуцируемых транскрипционных факторов, были названы, по аналогии с вирусными системами, "поздними" генами, "генами позднего ответа" или "эффекторными" генами, а весь двухфазный механизм регуляции транскрипции с участием этих двух классов генов является одним из наиболее универсальных способов обеспечения процессов клеточного деления и роста в развитии.
В середине 1980-х годов несколько исследовательских групп обнаружили экспрессию гена c-fos в мозге обучающихся взрослых животных. Это повлекло за собой широкий спектр исследований участия этого и других НРГ в самых разных задачах обучения.