Письмо, чтение и счет
Письмо
Все задания выполняются и правой и левой рукой.
1. Написание отдельных букв и слогов. Списывание и написание слов, упроченных в опыте: собственное имя, мама, домашняя работа и т.д.
2. Написание отдельных слов и словосочетаний: машина, Мишина машина; гвоздь, кораблекрушение, гвоздь—кость, гвоздь—грусть —гость, бочка —почка, почка —почта, бочка—почка —дочка» и т. п. Сначала задания выполняются в свободном режиме, потом с зафиксированным языком.
3. Написание предложений: «Портной строчит строчку», «Устроили экскурсию в Псков», «Лавировали корабли, пока не вылавировали».
Чтение
1. Прочтение простых и наложенных букв и цифр в «Альбоме» (с. 27).
2. Прочтение слогов, высокои малочастотных слов, неверно написанных слов и чисел (с. 28).
3. Прочтение рассказа (с. 47 — 49).
Счет
Исследование счета по существу уже описано выше в разных разделах. Это зрительный и пространственный цифровой гнозис, написание и чтение отдельных цифр и чисел, тест Шульте, воспроизведение числового ряда в прямом и обратном порядке; серийный счет «100 — 7» и «30— 1 и 2».
Заболевания головного мозга
Травматические повреждения головного мозга могут иметь разную степень тяжести: начиная от сотрясения головного мозга легкой степени тяжести, заканчивая ушибом мозга или его проникающим ранением, сопряженным с трепанацией черепа. Симптомы зависят от степени тяжести пострадавшего и диагноза. К примеру, при сотрясении головного мозга возникают кратковременная потеря сознания, затем тошнота и рвота, головокружение, сильная головная боль, слабость. При более тяжелом поражении возможны длительная потеря сознания и кома, серьезное угнетение рефлексов, судороги. В ряде случаев последствием тяжелой травмы головного мозга становится посттравматическая эпилепсия. Если произошло проникающее ранение или развивается прогрессирующая внутричерепная гематома, абсолютно показано оперативное вмешательство. Инфекционное поражение головного мозга возникает при вирусных и бактериальных инфекциях. Если воспалены оболочки мозга — это менингит, если в процесс вовлечено вещество мозга — это энцефалит, при сочетанном поражении ставят диагноз «менингоэнцефалит». Вирусные процессы чаще протекают более доброкачественно, нежели бактериальные. К примеру, поражение мозга при энтеровирусной инфекции в большинстве случаев достаточно быстро проходит без тяжелых последствий. С другой стороны, коревой, ветряночный и герпетический энцефалиты иногда способны давать злокачественное течение, приводящее к декортикации (полному отмиранию коры головного мозга), если больной долго находился в коме. Исходами менингококкового или туберкулезного менингита могут быть как полное выздоровление, так и инвалидизация и даже смерть больного. Сосудистая патология головного мозга достаточно многообразна. Она включает ухудшение кровоснабжения какого-либо участка головного мозга (допустим, при шейном остеохондрозе или стенозе кровоснабжающей артерии), аневризмы (локальное патологическое расширение кровеносного сосуда) и сосудистые мальформации, нарушение целостности артерии, приводящее к инсульту (кровоизлиянию в головной мозг). Доброкачественные опухоли головного мозга представляют меньшую опасность, чем рак вследствие того, что не прорастают в другие ткани и не дают метастазов (дочерних опухолей в других местах). Однако они медленно увеличиваются в размерах, оказывают давление на прилежащие мозговые структуры, вследствие чего в части случаев требуется оперативное лечение. Онкологическое заболевание головного мозга может быть первичным (если опухоль изначально развилась в головном мозге) или вторичным (когда первичный очаг расположен в другом органе, а головной мозг поражен метастазами). К этой категории относятся такие опухоли, как глиома, менингиома, невринома и другие. Подобные опухоли отличаются быстрым ростом, способностью прорастать в прилежащие ткани и давать метастазы (очаги опухоли, расположенные в других местах.) Другие виды патологии головного мозга встречаются реже и включают врожденные, дегенеративные заболевания и другие. Диагностику при подозрении на заболевание головного мозга проводят с учетом других симптомов, сопровождающих заболевание. Сначала проводят врачебный осмотр. Уже на этом этапе можно выявить признаки менингита или менингизма (положительные симптомы Кернига и Брудзинского). Серьезным подспорьем служит спинномозговая пункция, позволяющая затем провести анализ ликвора и выявить в нем наличие крови или болезнетворных микроорганизмов. Широко используют визуализирующие методы, такие как УЗИ, КТ или МРТ(последнее исследование позволяет детально рассмотреть любой участок головного мозга и точно установить диагноз).
Цитоархитектоника (расположение клеток) Мультиполярные нейроны коры головного мозга весьма разнообразны по форме. Среди них можно выделить: пирамидные, звёздчатые, веретенообразные, паукообразные, горизонтальные. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоёв: Молекулярный , Наружный зернистый, Пирамидальных нейронов, Внутренний зернистый , Ганглионарный (слой клеток Беца), Слой мультиформных (полиморфных) клеток. Кора полушарий головного мозга также содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции.На медиальной и нижней поверхности полушарий сохранились участки старой, древней коры, которые имеют двухслойное и трехслойное строение.Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их аксоныпроходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Основная масса волокон этого сплетения представлена ветвлениями дендритов нейронов нижележащих слоёв.Наружный зернистый слой образован мелкими нейронами , имеющими округлую, угловатую и пирамидальную форму, и звёздчатыми нейронами. Дендриты этих клеток поднимаются в молекулярный слой. Аксоны или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя.Слой пирамидальных нейронов Является самым широким по сравнению с другиями слоями коры головного мозга. Он особенно хорошо развит в прецентральной извилине. Величина пирамидных клеток последовательно увеличивается от наружной зоны этого слоя к внутренней. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Дендриты, берущие начало от боковых поверхностей пирамиды и её основания, имеют незначительную длину и образуют синапсы со смежными клетками этого слоя. Аксон пирамидной клетки всегда отходит от её основания. В мелких клетках он остаётся в пределах коры; аксон же, принадлежащий крупной пирамиде, обычно формирует миелиновоеассоцативное или комиссуральное волокно, идущее в белое вещество.Внутренний зернистый слойВ некоторых полях коры развит очень сильно (например, в зрительной зоне коры). Однако в других участках он может отсутствовать (в прецентральной извилине). Этот слой образован мелкими звёздчатыми нейронами. В его состав входит большое количество горизонтальных волокон.Ганглионарный слой (Внутренний пирамидный слой;Клетки Беца)Образован крупными пирамидными клетками, причём область прецентральной извилины содержит гигантские клетки, описанные впервые киевским анатомом В. А. Бецем в 1874 году (клетки Беца). Они достигают в высоту 120 и в ширину 80 мкм. В отличие от других пирамидных клеток коры гигантские клетки Беца характеризуются наличием крупных глыбок хроматофильного вещества. Их аксоны образуют главную часть кортико-спинальных и кортико-нуклеарных путей и оканчиваются на мотонейронахмозгового ствола и спинного мозга.Слой мультиморфных клетокОбразован нейронами различной, преимущественно веретенообразной формы. Внешняя зона этого слоя содержит более крупные клетки. Нейроны внутренней зоны мельче и лежат на большом расстоянии друг от друга. Аксоны клеток полиморфного слоя уходят в белое вещество в составе эфферентных путей головного мозга. Дендриты достигают молекулярного слоя коры.
Миелоархитектоника (расположение волокон)Среди нервных волокон коры полушарий головного мозга можно выделить:ассоциативные волокна — связывают отдельные участки коры одного полушария,комиссуральные волокна — соединяют кору двух полушарий,проекционные волокна — соединяют кору с ядрами низших отделов центральной нервной системы. Афферентные проекционные волокна заканчиваются в слое пирамидальных нейронов. модуль — вертикальная колонка.
ТРИ ИСТОЧНИКА ЗНАНИЙ О ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ МОЗГА
Наши знания о функциональной организации мозга животных и человека являются результатом использования трех следующих методических процедур:
во-первых, сравнительно-анатомических наблюдений;
во-вторых, физиологического метода раздражений отдельных участков мозга;
в-третьих, метода разрушения ограниченных участков мозга, а при исследовании функциональной организации мозга человека — клинических наблюдений над изменением поведения больных с локальными поражениями мозга.
1 СРАВНИТЕЛЬНО-АНАТОМИЧЕСКИЕ ДАННЫЕ
Изучение строения нервной системы — основного аппарата связи животного с внешним миром и регуляции его поведения — дает неоценимый материал для анализа того, что является субстратом психической деятельности на отдельных этапах психического развития, как осуществлялась регуляция поведения на последовательных этапах эволюции и что отличает нервную систему животных, живущих в различных условиях внешней среды и характеризующихся различными формами поведения.
Тесная связь строения нервного аппарата с уровнем организации поведения и экологическими особенностями животного позволяет широко использовать сравнительно-анатомический анализ для исследования способов жизни, особенностей поведения и основных принципов организации деятельности животных. Рассматривая строение нервной системы на последовательных этапах эволюции животного мира, можно выделить основные принципы этой эволюции. Основной и наиболее общий принцип заключается в том, что на различных этапах эволюции отношения организма животного со средой и его поведение регулировались различными аппаратами нервной системы, и, следовательно, мозг человека является продуктом длительного исторического развития. Хорошо известно, что на наиболее элементарных уровнях развития животного мира (например, у гидроидных полипов) прием сигналов и организация движений осуществляются диффузной, или сетевидной, нервной системой; на этом этапе эволюции единый центр, перерабатывающий информацию и регулирующий поведение животного, отсутствует и поток возбуждения определяется теми временными доминирующими очагами, которые создаются в том или ином участке нервного аппарата животного. Именно поэтому здесь можно говорить о временно доминирующих участках или органах, соответствующих этим временно наиболее возбудимым сегментам организма (Бете, 1931 и др.). В процессе эволюции диффузная сетевидная система (сохранившаяся в организме животных) уступила ведущее место новым образованиям. В передних отделах головного мозга животного концентрировались сложные рецепторные приборы, и сигналы, получаемые ими, стали направляться в передний ганглий, который перерабатывал получаемую информацию и переключал возбуждение на эфферентные пути, идущие к двигательному аппарату животного. На ранних ступенях эволюции (например, у червей) передний ганглий имел относительно простую функциональную структуру. На позднейших ступенях (например, у насекомых) по мере дифференциации системы рецепторов передний ганглий приобретает все более сложную функциональную организацию: в нем выделяются нейроны, изолированно реагирующие на обонятельные, зрительные и хемо-тактильные раздражения, промежуточные, ассоциативные нейроны и нейроны с двигательными функциями. Передний ганглий насекомых (например, пчел) становится идеальным органом реализации врожденного (инстинктивного) поведения, которое может пускаться в ход элементарными стимулами и тем не менее иметь удивительную по своей сложности программу. Нервные аппараты переднего ганглия, хорошо приспособленные для реализации врожденных программ поведения, не могут, однако, обеспечить приспособления к резко меняющимся условиям среды. В таких случаях сохранение вида оказывается возможным либо благодаря избыточному производству индивидуальных особей, из которых выживают лишь очень немногие, либо благодаря выработке индивидуально-изменчивого поведения. По второй линии идет развитие позвоночных. Если у низших позвоночных еще остается старый принцип сохранения индивида и вида, который хорош для существования в условиях однородной водной среды, то при переходе к наземному существованию появляется необходимость в нервных аппаратах, которые в отличие от переднего ганглия обеспечивали бы максимальную индивидуальную изменчивость поведения, соответствующую большой изменчивости условий жизни на земле. Таким биологическим задачам отвечает головной мозг. На ранних этапах эволюции позвоночных (например, у рыб и земноводных) он допускает лишь относительно небольшую изменчивость поведения. Преобладающие формы поведения реализуются здесь аппаратами элементарного обонятельного и среднего мозга (у рыб они являются единственными и поэтому ведущими нервными образованиями). С дальнейшим развитием к ним присоединяются нервные аппараты, позволяющие животному осуществить более сложные формы анализа и приспособления к условиям среды; у птиц ведущее место занимают уже аппараты межуточного мозга (зрительный бугор, подкорковые двигательные узлы), которые образуют таламо-стриальную систему, обеспечивающую более высокий уровень поведения, названный Н.А.Бернштейном (1947) уровнем синергий. У млекопитающих аппараты таламо-стриальной системы уступают ведущее место более сложным нервным аппаратам коры головного мозга, лежащим в основе огромного разнообразия форм индивидуально-изменчивого поведения. Корковые аппараты в полной мере способны получать и анализировать информацию, поступающую из внешней среды, перерабатывать ее, формировать новые связи и хранить их следы. Они в состоянии заменять врожденные программы поведения сложными индивидуально-изменчивыми, обеспечивая не только выработку условных рефлексов, но и формирование значительно более сложных программ индивидуального поведения. По мере эволюции высших позвоночных значение этих аппаратов все более возрастает, и на этапе человека, когда к естественным условиям среды прибавляются условия общественно-исторические и когда возникает язык — уникальная для человека система кодов, эти аппараты достигают такого уровня развития, что оказываются в состоянии обеспечить формы поведения, по степени сложности не имеющие равных в животном мире. Исследователи неоднократно пытались продемонстрировать прогрессивное развитие мозга на последовательных ступенях эволюции, принимая за показатель этого развития изменение отношения массы мозга к массе тела. Пожалуй, наиболее отчетливо это увеличение описывается индексом Хауга (1958):
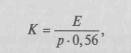
где Е — масса мозга, р — масса тела, а 0,56 — эмпирически найденный индекс; другими используется индекс Я.Я. Рогинского:
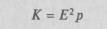
Цифры показывают, что если на каждое волокно зрительного нерва у крысы приходится лишь 10 нервных клеток коры, то у макаки их число увеличивается до 145, а у человека — до 500; аналогичное увеличение числа нейронов коры, приходящихся на одно нервное волокно, отмечается и в слуховой сфере (у обезьяны — преимущественно зрительного животного — увеличение не столь заметно); тот же принцип сохраняется и в отношении соответствующих отделов подкорки. Мы видим, таким образом, что в процессе эволюции удельный вес коры — по сравнению с нижележащими подкорковыми образованиями — непрерывно возрастает. Естественно, что большие полушария головного мозга и его кора становятся у человека важнейшим аппаратом регуляции поведения. Существенным является тот факт, что это огромное увеличение объема и массы мозга связано не с ростом наиболее древних, стволовых, отделов мозга, а в первую очередь с развитием больших полушарий и их наиболее существенной части — коры (рис. 3). Было бы неправильным думать, что все области коры человеческого мозга развиваются в процессе эволюции равномерно. Внимательный анализ показал, что развитие больших полушарий связано прежде всего с ростом новых областей коры, которые у низших млекопитающих едва намечены, а у человека составляют основную часть коры (табл. 3); древние области коры — палеокортекс (включающий образования коры, еще не отделенные от подкорковых образований), архикортекс (образования двуслойной древней коры, входящей в систему обонятельного мозга) и межуточная кора (образования, носящие переходный характер между только что упомянутыми), напротив, у человека составляют лишь незначительную часть коры, в то время как у низших млекопитающих они доминируют. С переходом от высших млекопитающих (обезьян) к человеку эволюция мозга связана преимущественно с увеличением площади наиболее сложных (третичных) зон коры; площадь более элементарных отделов коры (первичных и вторичных) практически не увеличивается (а иногда даже становится меньше) Эти данные убедительно показывают принципиальные изменения в соотношении отдельных зон мозговой коры при переходе от низших обезьян к высшим, а затем к человеку. Они позволяют видеть, что относительный размер более просто организованной лимбической области снижается с переходом к человеку; относительный размер прецентральной (двигательной) коры остается без изменений; размер первичной (проекционной) затылочной коры у человека даже уменьшается по сравнению с таковой у обезьяны, в жизни которой зрительное восприятие играет особенно большую роль. Напротив, размеры височной области у человека значительно увеличиваются, а размеры третичных полей коры — нижнетеменной и лобной областей — возрастают в несколько раз.
Мы видим, таким образом, какое огромное место в коре мозга человека отводится аппаратам, связанным с приемом, переработкой (кодированием) и синтезом информации, получаемой от различных анализаторов, и аппаратам, принимающим участие в выработке и сохранении сложнейших программ поведения и контроля психической деятельности.
Было бы, однако, глубоко неправильным думать, что если у человека кора головного мозга приобретает ведущую роль, то все нервные образования, которые на более низких этапах эволюции были единственными аппаратами, обеспечивающими организацию поведения, теперь совершенно отстраняются от работы. Важнейший принцип работы мозга заключается в том, что прежние нервные аппараты сохраняются в нем, но сохраняются, если пользоваться выражением Гегеля, в снятом виде, иначе говоря, сохраняются, уступая ведущее место новым образованиям и приобретая иную роль. Они все больше и больше становятся аппаратами, обеспечивающими фон поведения, принимающими активное участие в регуляции состояний организма, передавая как функции получения, переработки и хранения информации, так и функции создания новых программ поведения и регуляции и контроля сознательной деятельности высшим аппаратам коры головного мозга (рис. 5).
Мы хорошо знаем сейчас, что разные по сложности формы поведения даже у человека могут осуществляться с помощью различных уровней нервной системы.
Каждый физиолог и невролог хорошо знает, что такие простейшие элементы поведения, как сегментарные рефлексы (например, коленный рефлекс, элементарные защитные рефлексы), осуществляются лишь механизмами спинного мозга, и у больного, у которою ранение полностью отделило аппараты спинного мозга от более высоких уровней, эти рефлексы могут сохраняться или даже усиливаться, хотя и не улавливаются сознанием.
Физиологам известно также, что такая сложнейшая врожденная форма поведения, как регуляция обменного равновесия (го-меостаза), обеспечиваемая дыханием, пищеварением и терморегуляцией, осуществляется посредством механизмов, заложенных в верхних отделах ствола (продолговатом мозге, гипоталамусе); при нарушении их соответствующие процессы расстраиваются; грубые поражения этих механизмов могут привести к нарушению «витальных функций» и смерти.
Кроме того, физиологи и неврологи знают, что еще более сложные формы поведения, предполагающие обеспечение тонуса, синергий и координацию, тесно связаны с работой межуточного мозга и подкорковых двигательных узлов (таламо-спинальной системы); поражение их, не вызывая нарушения сложных познавательных процессов, приводит к грубому нарушению «фонового» поведения. Особый интерес в связи с этой проблемой представляют результаты наблюдений над больными, страдающими паркинсонизмом, накопившиеся за последние три десятилетия в результате широкого изучения эпидемического энцефалита и распространения стереотактических операций.
Наконец, хорошо известно, что наиболее сложные формы деятельности не могут быть обеспечены без участия коры головного мозга, являющейся органом высших форм поведения животных и сознательного поведения человека.
Таким образом, ясно, что сложные рефлекторные процессы и сложные формы поведения могут осуществляться разными уровнями нервной системы, каждый из которых вносит в функциональную организацию поведения свой вклад.
Последние десятилетия позволили во многом уточнить только что обозначенное положение. Было показано, что низшие уровни нервного аппарата участвуют в организации работы коры больших полушарий, регулируя и обеспечивая ее тонус.
Аппараты стволового уровня не работают в полной изоляции от коры головного мозга и сами испытывают ее регулирующее влияние.
Данные, полученные в современных анатомических и физиологических исследованиях, позволяют сформулировать принцип вертикального строения функциональных систем мозга, иначе говоря, принцип, согласно которому каждая форма поведения обеспечивается совместной работой разных уровней нервного аппарата, связанных друг с другом как восходящими (летальными), так и нисходящими (фугальными) связями, превращающими мозг в саморегулирующуюся систему.
Этот прочно вошедший в науку принцип утверждает, что кора головного мозга, находящаяся в постоянном взаимодействии с нижележащими образованиями, не является единственным мозговым субстратом психических процессов.
Этот принцип делает понятным и те факты, которые ставили в тупик многих исследователей прошлого. Было показано, что разобщение отдельных зон коры путем круговой изоляции может не влечь за собой существенных изменений в поведении животных, в то время как подрезка коры, изолирующая ее от нижележащих образований, неизбежно приводит к значительным нарушениям ее регулирующих функций (Чоу, 1954; Сперри, 1959; Прибрам, Блейрт, Спинелли, 1966). Все это означает, что отдельные участки коры головного мозга соединяются между собой не только с помощью горизонтальных (транскортикальных) связей, но и через нижележащие образования, иначе говоря, посредством системы вертикальных связей.
Имея в виду сформулированные выше положения об эволюции нервных аппаратов, основных уровнях нервной системы и их взаимодействии, мы переходим сейчас к рассмотрению данных, которыми располагает сравнительная анатомия коры головного мозга.
Структурная и функциональная организация коры головного мозга
Наблюдения, показавшие, что мозг в целом и его кора в частности обладают неоднородным строением, относятся еще к началу прошлого века.
Ф. Галль, известный анатом, вошедший в историю науки как основатель фантастической «френологии» (концепции о функциональной организации мозга, исходящей из представлений о локализации сложных психических «способностей» в его ограниченных участках), впервые отличил серое вещество, составляющее мозговую кору и подкорковые серые образования, от белого вещества, состоящего из проводящих волокон, связывающих отдельные участки коры и соединяющие кору большого мозга с периферией. Однако это открытие, сделавшее Галля подлинным основателем морфологии мозга, долго оставалось без адекватной оценки, и настоящее раскрытие функций коры головного мозга, ее проводящих путей и серого вещества, заложенного в глубине больших полушарий, было сделано лишь спустя несколько поколений.
Значительный шаг вперед был сделан в 1863 г. киевским анатомом В. А. Бецом, занимавшимся микроскопическим изучением клеточного состава мозговой коры. Ему принадлежит открытие, которому было суждено стать началом целой эпохи блестящих исследований.
Описывая строение различных участков мозговой коры, он обнаружил, что их морфологическая структура в высокой степени неоднородна: если кора передней центральной извилины включает в свой состав большие, имеющие форму пирамиды нервные клетки (они получили в дальнейшем название гигантских пирамидных клеток Беца), то прилегающая к ней кора задней центральной извилины имеет совсем иное, мелкозернистое строение и совсем лишена пирамидных клеток (рис. 6).
Позднее было установлено, что различие этих двух областей коры не только морфологическое, но и функциональное. Гигантские пирамидные клетки Беца (составляющие пятый слой коры) оказались источниками двигательных импульсов, идущих от коры к периферической мускулатуре, а передняя центральная извилина, в которой они были сосредоточены, — моторной областью коры головного мозга. Поля мозговой коры, имеющие мелкозернистое строение и отличающиеся развитым четвертым слоем нервных клеток (к их числу относятся и образования задней центральной извилины), оказались аппаратами, к которым подходят чувствительные волокна, начинающиеся в периферических органах чувств (рецепторах), а соответствующие зоны коры — первичными чувствительными образованиями коры большого мозга.
С выделением двигательных и сенсорных областей (или первичных двигательных и сенсорных центров) был сделан первый шаг к созданию функциональной карты коры головного мозга, и кажущаяся однородной масса серого вещества, покрывающая тонким слоем большие полушария, начала приобретать дифференцированный характер.
Дальнейшие сравнительно-анатомические наблюдения подтвердили плодотворность наметившегося подхода. Оказалось, что внимательное изучение «первичных» областей мозговой коры позволяет делать точные выводы о некоторых особенностях поведения животного. Следующие примеры хорошо иллюстрируют это положение.
На рисунке 7 мы приводим срезы из двигательных областей мозговой коры австралийского медвежонка (рис. 7, а) и летучей собаки (рис. 7, б). Легко увидеть, что на первом срезе присутствует сравнительно немного гигантских пирамидных клеток, в то время как на втором срезе число их значительно больше, а величина гораздо меньше. Не указывает ли нам этот факт на относительно сильные и грубые движения первого животного и более тонкие и многообразные движения второго?
К аналогичным заключениям позволяет прийти и сравнительно-анатомический анализ строения сенсорных отделов коры.
). Он указывает на ведущую роль обонятельного анализатора у низших млекопитающих и незначительное место этого анализатора у человека.
Подобный вывод можно сделать и при сравнении зрительных полей животных и человека (рис. 9): значительное развитие мелкоклеточных образований зрительной коры обезьяны, составляющих у нее до 40 % площади коры (рис. 9, б), по сравнению с такими же образованиями зрительной коры крота (рис. 9, а) объясняется тем, что ведущее место в поведении первого животного занимает зрение, в то время как в поведении второго животного, ориентирующегося в окружающем мире с помощью обоняния, оно занимает лишь сравнительно небольшое место.
Сравнительно-анатомическое изучение коры головного мозга, начавшееся с выделения основных, первичных, или проекционных, зон мозговой коры, существенно продвинулось за последние десятилетия.
Решающие успехи были сделаны еще в начале этого столетия, когда работы Кэмпбелла (1905) и Бродмана (1909), Рамон-и-Кахала (1909 — 1911) и Болтона (1933) позволили составить цитоархитектонические карты мозговой коры; эти карты, существенно уточненные Ц. и О. Фогтами (1919 — 1920) и работами Московского института мозга, позволили приблизиться к описанию основных принципов строения мозговой коры животных и человека и внесли неоценимый вклад в наши знания о мозге как органе психики.
Как показали эти исследования, новая кора головного мозга состоит из шести слоев клеток (рис. 10). Только нижние из них являются аппаратами, непосредственно связывающими мозговую кору с периферией: органами чувств (IV — афферентный слой) и мышцами (V — эфферентный слой). В IV слое коры приходят волокна, несущие импульсы, возникающие в периферических ре-
цепторах; именно этот слой мелкозернистых клеток особенно мощно развит в только что упомянутых первичных чувствительных зонах коры. V слой включает в себя гигантские пирамидные клетки, генерирующие импульсы к мышцам тела и дающие начало длинному двигательному пути, состоящему из нервных волокон; абсолютное преобладание этого слоя имеет место в передней центральной извилине, или двигательной зоне, коры головного мозга.
На рисунке 11 мы приводим схему, позволяющую проследить ход волокон от периферических органов чувств в соответствующие «проекционные» отделы коры головного мозга. Она показывает, что волокна, начинающиеся от чувствительных аппаратов кожи и мышц, прерываясь в подкорковых образованиях, приходят к коре задней центральной извилины (общечувствительная зона), а волокна, идущие от сетчатки глаза и от внутреннего уха, также переключаясь в подкорковых аппаратах, заканчиваются соответственно в затылочных и в первичных височных отделах коры.
Таким образом, в коре головного мозга человека выделяются проекционная общечувствительная (теменная), зрительная (затылочная) и слуховая (височная) области.
Аналогичным образом мы можем проследить волокна, которые, начинаясь в передней центральной извилине и подходя к передним рогам спинного мозга, несут двигательные импульсы к мышцам. Эти волокна составляют двигательный, или пирамидный, путь головного мозга.
Как показали морфологические исследования, над каждой «первичной» зоной коры (с преобладающим развитием IV — афферентного или V — эфферентного слоев клеток) надстраивается система «вторичных» зон, в которых преобладающее место занимают более сложные по своему строению II и III слои. Эти слои состоят из клеток с короткими аксонами, большая часть которых или не имеет прямой связи с периферией, или получает свои импульсы из лежащих в глубине мозга подкорковых образований, осуществляющих первичную переработку приходящих с периферии импульсов. Строение этих слоев позволяет относить их уже не к простейшему — «проекционному», а к гораздо более сложному — «ассоциативному», или «интегрирующему», аппарату коры головного мозга.
Существенным для понимания функции этих слоев коры головного мозга является тот факт, что в процессе эволюции видов удельный вес их непрерывно увеличивается (рис. 12); это показывает, что процесс усложнения психической деятельности, переход от относительно простых, врожденных форм поведения животного к более сложным формам кодирования поступающей информации у человека, предполагающим сознательный характер программирования деятельности, связаны с развитием этих высших слоев мозговой коры.
Другой не менее важной функциональной характеристикой строения коры мозга животного является отношение между массой клеточных тел и массой клеточного вещества.
Исследования последнего времени показали, что в осуществлении сложных нервных процессов решающую роль играет не только тело нервной клетки, но и ее многочисленные отростки и, наконец, окружающие нейроны глиальные клетки (Хиден, 1962, 1964; Ройтбак, 1965; и др.).
Увеличение «глиального индекса» на каждой новой ступени эволюции указывает на повышение управляемости функций отдельных мозговых зон, однако лишь будущие сравнительно-анатомические исследования могут выявить его недлинный функциональный смысл.
Характерным поэтому является тот факт, что с эволюцией животного величина отношения глиальной ткани коры к массе ее нервных клеток все более возрастает и у человека оказывается во много раз большей, чем у млекопитающих, стоящих на более низких ступенях эволюции (табл. 5).
Аналогичная тенденция легко прослеживается в процессе созревания коры мозга человека. У плода 6 месяцев верхние слои коры едва намечены, у младенца — развиты относительно слабо, у нормального взрослого — занимают значительное место (рис. 13). В случаях врожденного слабоумия эти слои клеток недоразвиты, а у больных с органической деменцией и атрофией коры — резко сужены.
Все это указывает на то, что верхние, «ассоциативные», слои мозговой коры играют важную роль в осуществлении наиболее сложных форм психической деятельности, становление которых происходит на поздних ступенях филогенеза и на поздних этапах развития человека.
Не менее важным анатомическим фактом, позволяющим нам понять основные принципы строения мозговой коры, является неравно-
мерность распределения отдельных слоев коры в топографически различных участках коры головного мозга.
Факты показывают, что над каждой первичной областью мозговой коры, в которой преобладают низшие (афферентные или эфферентные) слои коры, надстраиваются вторичные области, в которых преобладают верхние (проекционно-ассоциационные) слои, играющие, как мы увидим далее, существенную роль в функциональной организации работы отдельных анализаторов.
Иерархическое строение мозговой коры легко можно видеть, рассмотрев топографическую карту мозга, изображенную на рисунке 14. Мы видим, что над первичными (проекционными) отделами общечувствительной коры (задняя центральная извилина) у человека надстраивается вторичная чувствительная кора, в которой преобладают верхние (проекционно-ассоциационные) слои; над первичной зрительной корой, расположенной в полюсе затылочной области, надстраивается вторичная зрительная кора, где также преобладают верхние (проекционно-ассоциационные) слои; над первичной слуховой корой, расположенной в верхних отделах височной области, надстраиваются ее вторичные отделы с тем же строением; наконец, над первичной двигательной корой, занимающей переднюю центральную извилину, — ее вторичные отделы, расположенные в премоторной области.
Как видно из той же карты, в коре головного мозга человека можно выделить участки, которые лежат на границах между корковыми представительствами отдельных чувствительных зон мозговой коры и которые получили название третичных зон коры (или зон перекрытия коркового представительства отдельных анализаторов). Эти области коры целиком состоят из верхних (ассоциационных) слоев клеток и не имеют прямой связи с периферией. Есть все основания предполагать, что третичные зоны коры обеспечивают совместную работу корковых звеньев отдельных анализаторов, наиболее сложные интегральные функции коры головного мозга.
Как показали детальные анатомические исследования, в коре головного мозга можно выделить две группы третичных областей. Первая из них — задняя — расположена на стыке зрительной (затылочной), общечувствительной (теменной) и слуховой (височной) областей; ее с полным основанием можно обозначить как зону перекрытия корковых отделов экстероцептивных анализаторов. Вторая — передняя — расположена спереди от двигательной зоны коры и надстраивается над двигательными отделами коры головного мозга. Она связана со всеми остальными отделами коры и, как мы увидим далее, играет существенную роль в построении наиболее сложных программ поведения человека.
Внимательное изучение хода волокон от периферических органов чувств к коре головного мозга в полной мере подтверждает принцип иерархического строения основных отделов мозговой коры. Рисунки 14 и 15 убедительно показывают, что функциональные системы головного мозга имеют одинаковое (иерархическое) строение и что анатомические данные позволяют выделить в коре головного мозга первичные, вторичные и третичные зоны.
Было бы неверным думать, что три описанных выше типа зон мозговой коры отмечаются на всех ступенях эволюции позвоночных. Факты говорят об обратном и указывают на то, что описанное нами иерархическое строение коры головного мозга является продуктом длительного исторического развития.
Как показывают сравнительно-анатомические данные, схематически представленные ранее на рисунке 4, в коре головного мозга ежа и крысы дифференциация первичных и вторичных зон едва намечается, а третичные зоны коры совсем отсутствуют; близкое строение имеет кора головного мозга собаки, лишь у обезьяны вторичные и третичные зоны мозговой коры отмечаются достаточно ясно. У человека иерархическое строение коры головного мозга выступает с полной отчетливостью; схема на рисунке 14 показывает, что первичные участки мозговой коры занимают у него совсем небольшое место, будучи оттесненными хорошо развитыми вторичными участками, и что третичные зоны мозговой теменно-височно-затылочной и лобной коры становятся здесь наиболее развитыми системами и занимают подавляющую часть коры больших полушарий.
В самое последнее время были получены очень важные данные, помогающие заполнить существенный пробел в наших знаниях о предыстории человеческого мозга.
Мы только что привели данные, касающиеся основных тенденций прогрессивного развития вторичных и третичных зон коры на последовательных этапах эволюции животного мира. Один вопрос остался, однако, неосвещенным: можно ли проследить такую же тенденцию и на последовательных этапах антропогенеза? Знаменуется ли переход от приматов к гоминидам и последующий переход от древних форм предчеловека к более поздним и совершенным формам соответствующим возрастанием удельного веса вторичных и третичных зон коры, отражающим усложнение условий жизни и переход от животной эволюции к социальной истории человека?
До сих пор мы знали лишь то, что объем головного мозга от высших обезьян к человеку постепенно растет и что с переходом к человеческой истории этот рост останавливается. По вычислениям Бонина (1934), средняя емкость черепа кроманьонца равнялась в среднем 1570 см3, приблизительно такая же емкость черепа характерна и для представителей неолита (1525 — 1533 см3), железного века (1514 см3) и различных периодов истории Древнего Египта (1390-1495 см3).
Означает ли это, что на всем протяжении антропогенеза соотношение отдельных образований мозговой коры оставалось неизменным?
До недавнего времени сколько-нибудь обоснованный ответ на этот вопрос казался невозможным. Только в последние годы были достигнуты первые успехи в решении этой проблемы. Связаны они с использованием оригинального метода получения слепков мозга с внутренней поверхности черепа (эндокраниума) различных предков человека. Наряду с учеными других стран большую работу в этом направлении проделала отечественный антрополог
В древнейшей истории человека можно выделить четыре больших этапа, представителями которых являются соответственно австралопитеки (или прегоминиды), жившие 1— 4 млн лет назад, архантропы (питекантропы, синантропы), жившие 200 тыс. — 1 млн лет назад, палеоантропы (неандертальцы, люди, найденные при раскопках в Ля Шапелле, Брокен-Хилле, Тешик-Таше), жившие 200 — 400 тыс. лет назад, и неоантропы (кроманьонцы), жившие 10 — 40 тыс. лет назад.
От австралопитеков остались лишь отдельные признаки предкультуры, архантропы и палеоантропы имеют четкие признаки шелльской, ранней и поздней ашёльской и мустьерской культуры; после неоантропов встречаются более богатые находки позднего палеолита. Охотничий образ жизни, требующий четкой ориентировки в пространстве, и употребление, а затем и производство элементарных орудий знаменовали первую фазу становления человека; на следующем этапе к нему, по-видимому, присоединяется возникновение языка, и именно эти два основных признака и составляют поворотный пункт, отделяющий естественную эволюцию животных от общественной истории человека.
Как показали сравнительные исследования В. И. Кочетковой, сводные результаты которых приводятся далее, соотношения частей мозга у представителей указанных выше четырех этапов эволюции существенно различны.
На рисунке 17 мы привели сравнительную серию фронтальных срезов и сводную схему горизонтальных срезов через слепки мозга — за основу сравнения в обоих случаях берется слепок мозга высшей обезьяны (шимпанзе), который последовательно сопоставляется со слепками мозга архантропов, палеоантропов и современных людей.
Нетрудно заметить, что слепок мозга антропоидной обезьяны имеет сферическую форму; отдельные части его не выступают 'отчетливо ни на фронтальных, ни на горизонтальных срезах.
Иное мы видим на слепках мозга ранних гоминид.
Уже на срезах через заднетеменную область (g. supramar-ginalis и g. angularis) (рис. 17, а, б) появляется выраженное местное увеличение мозга, заметное у архантропа и особенно резко выступающее у палеоантропа; аналогичную картину можно видеть на срезах через височную область (рис. 17, в), которая особенно мощно разрастается у палеоантропов. Исключительный интерес представляют срезы через заднелобную область (рис. 17, г), на анализе которых следует остановиться особо. Как показывают данные, нижнелобная область начинает существенно развиваться уже у синантропов и достигает значительного развития у палеоантропов; однако верхние отделы префронтальной области у синантропов по-
чти не развиты, несколько в большей степени они представлены у палеоантропов и только у современного человека получают наибольшее развитие. Подобную тенденцию можно увидеть при сопоставлении горизонтальных срезов (рис. 17, д).
Приведенные данные показывают, что основное развитие мозга в раннем антропогенезе идет не столько за счет наиболее элементарных (первичных), сколько за счет более сложных (вторичных и третичных) областей коры. Особый интерес для нас представляет последовательность развития отдельных областей больших полушарий. Оказалось, что из сложных областей коры в первую очередь начинают развиваться нижнетеменные области, значительный скачок в развитии которых отмечается уже у архантропов (синантропов и питекантропов), где они достигают существенных размеров. Есть все основания думать, что условия охотничьей жизни, требующие высокоразвитой способности ориентироваться в пространстве, составляют основу этого развития.
Во вторую очередь развиваются нижнелобные области мозга, которые достигают относительно больших размеров у палеоантропов. Это связано с возникновением и развитием звукового языка, который, как мы увидим далее, опирается на совместную работу височных и нижнелобных долей мозга.
Верхние отделы переднелобной области, как это отчетливо видно на рисунке 17, г, остаются относительно малоразвитыми на всех перечисленных выше этапах исторического развития прачеловека и интенсивно развиваются лишь у неоантропа и у современного человека. На рисунке 18 мы даем сводные данные, из которых видно, что у архантропов зона наиболее интенсивного роста расположена в нижнетеменной, у палеоантропов — в нижнелобной и у современного человека — в верхнелобной областях больших полушарий.
Далее мы собираемся более подробно говорить об описанных здесь фактах, а также о той роли, которую играют указанные нами области больших полушарий в организации сложнейших форм сознательной деятельности. Отметим здесь только, что все приведенные данные отчетливо указывают на прямую связь эволюции головного мозга с усложнением процесса переработки и кодирования информации, с одной стороны, и с усложнением программ индивидуально-изменчивого поведения, которые отличают деятельность высших животных, — с другой.
Эти данные показывают также решающее значение эволюционного анализа анатомических изменений мозга для понимания процесса формирования сложных видов психической деятельности современного человека.
Большой интерес в плане темы данной книги представляет анализ тех изменений в мозговых структурах, которые наблюдаются в процессе развития ребенка.
Как показали исследования, ребенок появляется на свет с полностью созревшими аппаратами подкорковых образований и наиболее простых, проекционных или первичных, зон коры и с недостаточно созревшими аппаратами более сложных вторичных и третичных зон коры; это проявляется в относительно малом размере входящих в их состав клеток, в недостаточном развитии ширины их верхних слоев (имеющих, как известно, сложные ассоциативные функции), в относительно малой площади занимаемых ими территорий и, наконец, в недостаточной миелинизации их элементов.
Признаки недоразвития высших (вторичных и третичных) полей коры головного мозга исчезают постепенно и неравномерно, иными словами, коэффициенты роста отдельных полей мозговой коры неодинаковы. Если прижизненные коэффициенты роста таких относительно более простых областей коры, как лимбическая область, кора островка и кора первичных отделов центральной, затылочной и верхневисочной областей, не выходят за пределы
2,5— 5 усл. единиц, то площадь наиболее сложных, вторичных и третичных, областей коры увеличивается более интенсивно — коэффициент роста 7 —9 усл. единиц (рис. 19). Согласно материалам Московского института мозга, наиболее бурное увеличение территории последних областей отмечается в возрасте 2 — 3 лет, причем наиболее сложные лобные области созревают окончательно лишь к 6 —7-летнему возрасту (рис. 20).
Аналогичным образом развиваются в онтогенезе верхние (ассоциативные) слои только что упомянутых зон коры. И здесь мы отмечаем уже знакомый нам факт особенно интенсивного роста ширины этих функционально наиболее важных слоев коры к 3 — 3,5 года жизни ребенка, причем увеличение их в случае некоторых особенно сложных полей продолжается до 7- и даже 12-летнего возраста. Этот факт отчетливо говорит о том, что по мере развития ребенка возрастает роль тех видов деятельности, которые требуют совместной работы отдельных зон коры и осуществляются при ближайшем участии верхних, ассоциативных, или интегративных, ее слоев.
Более подробное описание только что указанной тенденции содержится в таблице 6, составленной на основании работ целой группы отечественных морфологов. Из таблицы видно, что если для наиболее простых (первичных, или проекционных) зон коры характерно незначительное развитие в онтогенезе, то более сложные — вторичные и третичные — зоны коры развиваются в онтогенезе весьма интенсивно.
Увеличение площади вторичных и третичных зон коры на сравнительно поздних этапах филогенеза является далеко не единственным признаком, позволяющим судить о их готовности к участию в регуляции поведения человека. Столь же существенной характеристикой является степень миелинизации соответствующих нервных образований.
Как показали классические исследования Флексига (1900, 1927), процесс миелинизации — по завершении которого нервные элементы становятся готовыми к нормальному функционированию — протекает в разных зонах коры неравномерно: если миелинизация элементов первичных (проекционных) зон заканчивается довольно рано, то процесс миелинизации во вторичных и третичных зонах коры затягивается на очень длительные сроки и в некоторых случаях продолжается до 7 —12-летнего возраста. Достаточно посмотреть на миелогенетическую карту, составленную Фогтами (рис. 21), на которой изображены зоны, элементы которых наиболее рано кончают миелинизацию (крупные кружки), зоны, которые миелинизируются в последнюю очередь (мелкие точки), и зоны, занимающие промежуточное место (средние точки), чтобы отчетливо увидеть, что аппараты, соответствующие наиболее сложным, комплексным формам психической деятельности созревают на относительно поздних этапах развития и что, следовательно, формирование психической деятельности человека идет от более простых к сложным, опосредствованным формам.
Далее, на протяжении книги, нам не один раз представится случай убедиться в том, насколько большое значение имеют полученные в эволюционной анатомии данные для решения важнейших проблем психологической науки.
ФИЗИОЛОГИЧЕСКИЕ ДАННЫЕ: МЕТОД РАЗДРАЖЕНИЯ
Опыты с непосредственным раздражением коры
Данные, полученные в сравнительно-анатомических исследованиях, составляют основу наших знаний о функциональной организации мозга как органа психической жизни; однако они не дают прямых указаний на нервные механизмы, связанные с приемом информации от внешнего мира и регуляцией поведения человека.
Существенный прогресс в этом направлении связан с применением метода раздражения отдельных участков мозга. Благодаря ему физиологи смогли получить сведения о непосредственной функции тех или иных мозговых систем.
Еще во второй половине XIX века исследователи раздражали отдельные пункты коры головного мозга животных, с тем чтобы выявить таким путем функции различных участков мозга. В 1871 г. были опубликованы результаты известных опытов Фрича и Гитцига, установивших, что раздражение электрическим током определенных участков коры головного мозга собаки вызывает сокращение мышц противоположных конечностей. Так была впервые выделена двигательная зона коры и положено начало точному физиологическому исследованию мозговых функций (рис. 22).
Спустя сравнительно небольшой период времени классические исследования, проведенные Чарльзом Шеррингтоном и его сотрудниками (Грюнбаум и Шеррингтон, 1903; Лейтон и Шеррингтон, 1917), показали, что двигательная зона коры головного мозга обезьяны имеет четкую функциональную организацию. Оказалось, что гигантские пирамидные клетки верхних отделов передней центральной извилины дают начало волокнам, идущим к нижним, а гигантские пирамидные клетки ее нижних отделов — к волокнам, идущим к верхним конечностям противоположной стороны тела. (Раздражение соответствующих пунктов двигательной зоны слабым электрическим током вызывало сокращение строго определенных мышц противоположной стороны тела.) Опыты Шеррингтона и его сотрудников положили начало изучению функциональной организации мозговой коры объективными физиологическими методами. Успехи нейрохирургии очень скоро сделали возможным дальнейший прогресс в этой области; через два десятилетия после первых публикаций Шеррингтона О. Ферстер, один из основоположников нейрохирургии, смог показать, что предложенные Шеррингтоном методические принципы пригодны и для анализа функций коры головного мозга человека. Последовавшие за исследованиями О. Ферстера работы Г. Хоффа и О.Петцля (1930) и, наконец, выдающегося канадского нейрохирурга В. Пенфилда и его сотрудников (Пенфилд и Эриксон, 1945; Пенфилд и Расмуссен, 1950; Пенфилд и Джаспер, 1959) создали прочную основу учения о функциональной организации различных зон мозговой коры человека.
Опыты Пенфилда не только подтвердили, что сформулированный Шеррингтоном применительно к мозгу обезьяны принцип функциональной организации моторной зоны коры верен и для мозга человека, в них были разработаны два принципиально новых аспекта исследования.
Во-первых, возможность наблюдать человека во время тонких нейрохирургических операций позволила расширить сферу изучения от двигательных функций коры до ее сенсорных функций: раздражая кору задней центральной извилины, а также затылочной и височной областей, исследователь мог спрашивать у больного об испытываемых им ощущениях и таким образом выявлять сенсорные функции этих зон коры. Таким образом, очень скоро оказалось возможным, не ограничиваясь исследованием функциональной организации двигательной сферы мозга, перейти к изучению функциональной организации афферентных, сенсорных систем.
Во-вторых, эти исследования позволили выйти за пределы простой констатации соответствия отдельных пунктов коры определенным участкам тела или, как это принято было говорить, принципа соматотопической организации первичных зон мозговой коры. Пенфилд показал, что как передняя, так и задняя центральные извилины организованы не столько по принципу линейной зависимости между размерами соответствующих участков тела и их проекциями на кору головного мозга, сколько по функциональному принципу: чем большее значение имеет та или другая функциональная система, тем более обширную территорию занимает ее проекция в первичных отделах коры 'головного мозга.
Оказалось, что проекции туловища, бедра или плеча занимают в передней и задней центральных извилинах относительно небольшую площадь; проекция руки — гораздо большую, а проекции губ и языка — еще большую площадь (рис. 23). Оказалось далее, что проекции 3-го и 4-го пальцев в коре сравнительно малы, тогда как участки, раздражая которые можно вызвать движения большого или указательного пальца, очень велики.
Так возникли известные схемы Пенфилда (рис. 24), наглядно показывающие, что чем более управляемым является тот или иной орган, тем шире он представлен в мозговой коре.
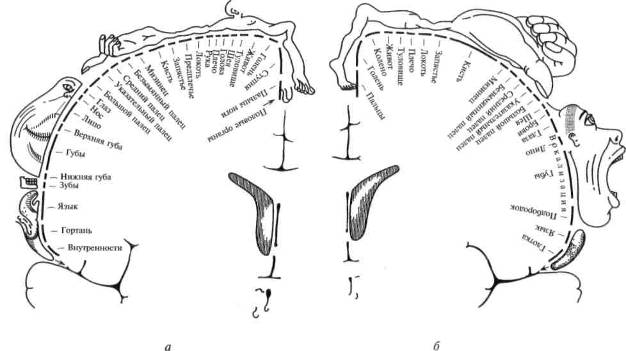
Рис. 24. Сравнительная площадь проекции отдельных поверхностей тела в проекционных зонах мозговой коры
(по Пенфилду):
а — корковая проекция чувствительности; 6 — корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения
Еще более углубили наши представления о физиологических механизмах первичных зон коры исследования, в которых было показано, что колонки клеточных элементов, соответствующие функционально наиболее важным системам органов, наиболее чувствительны к раздражениям и что существует «дифференциальная реактивность» (Differential accessibility) систем нервного аппарата, имеющих различное функциональное значение (Маунт-кастл, 1966; Филлипс, 1956, 1966). В этом отношении важны также многочисленные данные, показывающие, что пороги возбудимости клеточных элементов могут меняться под влиянием воздействий со стороны нижележащих отделов мозга (Мэгун и Моруцци, 1949; Линдсли и др., 1949; Френч, 1952; Джаспер, 1963, 1966; Эрнандес-Пеон, 1966; и др.).
Применение описанных физиологических приемов исследования функциональной организации мозга не ограничилось опытами с раздражением первичных зон мозговой коры. Результаты, значение которых для понимания функциональной организации коры головного мозга трудно переоценить, были получены в экспериментах со стимуляцией вторичных отделов мозговой коры, отличающихся, как было указано ранее, мощным развитием верхних (ассоциативных) слоев (рис. 25).
Едва ли не самые интересные результаты были получены американским исследователем Мак-Кэллоком (1943, 1944), сумевшим ответить на вопрос о том, как далеко распространяется возбуждение, вызываемое раздражением определенного пункта коры головного мозга. Предложенный автором нейронографический метод был очень прост: небольшую бумажку, смоченную раствором стрихнина, налагали на определенный участок коры головного мозга, тем самым раздражая его. Затем электроды последовательно прикладывали к соседним участкам, «прощупывая» таким образом, далеко ли распространяется вызванное раздражение.
Эксперимент показал, что если раздражение первичных зон коры распространялось лишь на области, непосредственно прилегающие к раздражаемому пункту, то возбуждение, вызванное раздражением вторичных зон коры, охватывало целый комплекс зон коры, иногда расположенных на значительном расстоянии от раздражаемого пункта (рис. 26). Этот факт свидетельствует о том, что в процесс возбуждения, возникающий во вторичных отделах коры, вовлекаются большие системы нервных элементов и, таким образом, обеспечиваются значительно более сложные, интегральные, процессы по сравнению с теми, которые имеют место при возбуждении первичных зон.
Значение такого обширного распространения возбуждения, исходящего из вторичных зон мозга, становится ясным из опытов, в которых эффект раздражения проверялся с помощью не электрофизиологических, а психологических показателей. Результаты этих опытов настолько важны, что на них следует остановиться особо.
Эксперименты с раздражением коры головного мозга больного на операционном столе, проведенные О.Петцлем, а затем Пенфилдом и его сотрудниками, показали, что эффект раздражения определенных пунктов первичных зон коры резко отличен от того, который вызывается раздражением вторичных зон. Это с особенной отчетливостью видно при раздражениях соответствующих отделов затылочной (зрительной) и височной (слуховой) коры.
Раздражение первичных отделов коры вызывало у больного элементарные ощущения. Так, больной, у которого на операционном столе раздражали полюс затылочной зоны, заявлял, что он внезапно начинает видеть мелькающие световые точки, окрашенные шары, языки пламени и т.п., причем эти неоформленные зрительные галлюцинации (фотопсии) располагались в различных участках зрительного поля в строгой зависимости от того,
какое место коры раздражалось (они появлялись в верхних секторах поля зрения, если раздражались нижние участки зрительной коры, и в нижних секторах, если раздражались верхние участки). Аналогичная картина наблюдалась при раздражении первичных участков височной (слуховой) коры, с той только разницей, что в этих случаях у человека появлялись элементарные слуховые галлюцинации (шумы, тоны).
Совершенно иными явлениями сопровождается раздражение вторичных отделов коры. Раздражение передних отделов затылочной области (вторичной зрительной коры) вызывает сложные оформленные зрительные образы: испытуемый видит людей, зверей, бабочек, человека, идущего или делающего знаки рукой, птиц в полете и т. п. Следовательно, возбуждение, вызванное раздражением этой области коры, не только распространяется на более обширные площади, но и приводит к всплыванию законченных зрительных образов, сформировавшихся у человека в прежнем опыте.
Аналогичные факты отмечаются при раздражении соответствующих отделов височной (слуховой) коры. Если, как уже было указано, раздражение первичных отделов слуховой коры (трудноосуществимое из-за того, что первичные зоны слуховой коры расположены в малодоступной поперечной извилине височной области) вызывает лишь ощущение тонов или шумов, то раздражение вторичных отделов слуховой коры, расположенных в верхней и средней извилинах височной области, приводит к появлению сложных слуховых галлюцинаций — музыкальных мелодий, а иногда (если раздражается височная кора левого полушария) к звучанию слов, фраз, песен, которые испытуемый слышит, полностью сознавая отсутствие их внешнего источника, так что они оказываются чем-то средним между реальными слуховыми ощущениями и слуховыми воспоминаниями.
Интересны опыты с раздражением еще более сложных отделов височной коры. Пенфилд, который провел большую серию таких исследований, мог наблюдать, что раздражение этих областей коры вызывает сложные сценоподобные галлюцинации — целые картины, иногда включающие в свой состав как зрительные, так и звуковые компоненты. Помимо опытов с раздражением определенных пунктов коры электрическим током аналогичные явления наблюдаются в тех случаях, когда рубец, образовавшийся в мозговой ткани (или оболочке мозга) после ранения или кровоизлияния, оказывает постоянное раздражающее воздействие на соответствующие участки мозга. Накопление такого раздражения может привести к эпилептическим припадкам (например, к так называемой Джексоновской эпилепсии), причем этим припадкам предшествуют явления, носящие характер зрительных или слуховых предвестников (ауры), сложность которых зависит от локализа-
ции рубца. Если рубец расположен в пределах первичных зон зрительной или слуховой коры, аура носит характер элементарных зрительных или слуховых галлюцинаций; если рубец расположен в более сложных — вторичных областях коры, она может принимать характер сложных, оформленных (зрительных или слуховых) галлюцинаций. Таким образом, характер ауры, предшествующей эпилептическому припадку, служит для невропатолога указанием на местоположение рубца, являющееся «эпилептогенной зоной».
Дополнение представлений об анатомическом строении отдельных зон мозговой коры данными, полученными при раздражении ее различных участков, позволяет значительно расширить наше понимание функциональной организации коры человеческого мозга.
Обе эти группы фактов подтверждают сформулированное ранее положение о том, что отдельные системы коры головного мозга имеют иерархическое строение и что возбуждение, возникающее в периферических органах чувств, сначала приходит в первичные (проекционные) зоны, отдельные участки которых представляют собой расположенные по топографическому принципу проекции соответствующих периферических рецепторов, и лишь вслед за этим распространяется на вторичные зоны коры, которые, опираясь на аппарат верхних (ассоциативных) слоев нейронов, играют интегрирующую роль, объединяя топографические (соматотопические) проекции возникших на периферии возбуждений в сложные функционально организованные системы.
По иерархическому принципу построены и двигательные отделы коры: над проекционными отделами передней центральной извилины (моторная зона) надстраиваются вторичные отделы премоторной области, располагающие мощно развитыми верхними (ассоциативными) слоями нейронов и способные превратить отдельные двигательные импульсы в целые системы возбуждений, составляющие физиологическую основу сложных предметных движений и двигательных навыков. Подробно о функциональной организации систем двигательной коры мы будем говорить далее.
Открытие системного характера построения основных зон мозговой коры является одним из главнейших этапов формирования наших представлений о функциональной организации человеческого мозга, и мы еще много раз сможем убедиться в важности этого фундаментального принципа.
Опыты с непрямой стимуляцией коры
До сих пор мы останавливались на результатах классических исследований, основанных на непосредственном электрическом или механическом раздражении коры головного мозга.
Существуют и другие, гораздо более естественные методы изучения функций головного мозга, которые, однако, настолько давно известны в психологии и физиологии, что мы упомянем о них здесь лишь в самых общих чертах. Некоторые из этих методов легли в основу объективного изучения поведения животного. Животному, например, предъявляли какой-нибудь стимул (обычно связанный с инстинктивной деятельностью и являющийся для него безусловным раздражителем), после чего изучали изменения в поведении животного, которые вызывались этим стимулом. И.П.Павлов предложил метод условных рефлексов, где условные раздражители (свет, звук, прикосновение), сами по себе способные вызвать лишь элементарную ориентировочную реакцию, в сочетании с безусловными раздражителями вызывают специфические — пищевые, оборонительные или половые — реакции. Этот метод открыл новые перспективы в изучении мозговой деятельности.
Работы Павлова настолько известны, что мы не будем останавливаться на них специально. Один из приемов «непрямой» стимуляции мозговой коры заслуживает, однако, того, чтобы на нем остановиться особо. Заключается этот прием в том, что раздражение адресуется не непосредственно коре, а периферическому отделу анализатора (коже, глазу, уху), после чего в отдельных участках коры головного мозга или в подкорковых образованиях прослеживается эффект такого раздражения. Один из наиболее важных вариантов этого метода называется методом вызванных потенциалов.