Сiмдiктiң қоректенуi, оның түрлері
Өсімдіктің сыртқы ортамен зат алмасуын оның қоректенуі дейді. Өсімдік қоректік заттарды жапырақтары және тамырлары арқылы қабылдайды. Осыған орай оның қоректенуінің екі түрі: ауа арқылы және тамыр арқылы қоректенуі болады. Өсімдік ауа арқылы қоректенуде өзінің жасыл жапырақтарының көмегімен, атмосферадағы көмірқышқыл газын ассимиляциялайды. Ал, тамыр арқылы қоректенуде топырақтағы суды, минералдық тұздардың иондарын және өте аз мөлшерде органикалық қосылыстарды сіңіреді. Соңғы жылдардағы зерттеулер органикалық заттар өсімдіктің тамырында түзіле бастайтындығын дәлелдеп отыр. Сондықтан өсімдіктің осы екі қоректену түрі бір-бірімен тығыз байланысты.
Өсімдіктің ауа арқылы қоректенуі. Өсімдік күн энергиясын жапырақтағы хлорофилл пигментінің қатысуымен сіңіріп, оны химиялық энергияға айналдырады. Нәтижесінде азотсыз органикалық зат (глюкоза) және оттек пайда болады. Жалпы алғанда фотосинтез реакциясының жиынтық теңдеуін былайша өрнектейді:
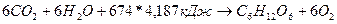
Өсімдік фотосинтез процесінің нәтижесінде түзілген оттектің шамалы бөлігін тыныс алуға пайдаланады, ал көбі атмосфераға ұшып кетеді. Глюкоза күрделі көмірсулар мен белок, май, органикалық қышқылдардың синтезделуіне қатысады. Фотосинтетикалық аппараттың қызметі өсімдік түріне, жапырақ жасына, күн сәулесінің құрамына, өсімдіктің азот элементімен қоректену деңгейіне т.б. жағдайларға байланысты. Егіншілік мәдениетін көтеруде фотосинтез процесінің мүмкіншілігін толық, әрі үнемді пайдаланудың маңызы үлкен. Өсімдікке жарықтың түсу дәрежесі фотосинтез процесінде аса маңызды рөл атқарады. Орташа есеппен жапырақ өзіне түсетін күн сәулесі спектрінің көрінетін бөлігіндегі энергияның 80-90% -ін және жарықтың жалпы энергиясының 50%-ін сіңіреді. Бүл сіңірілген энергияның 90%-і траспирацияға және жапырақты қыздыруға, ал небәрі 1-5%-тін фотосинтез процесіне жұмсалады. Фотоситезге хлорофилл сіңіретін қызыл және күлгін - көк сәулелер пайдаланылады. Бүл сәулелерді физиологиялық активті (белсенді) радиация - ФАР деп атайды. Жарық сапасы фотосинтездің қарқынына ғана емес, сондай-ақ түзілетін өнімнің химиялық табиғатына да әсер етеді.
Жалпы өсімдік құрамындағы заттарды талдағанда олардың 90-95%-ті фотосинтез процесінің арқасында түзілетіні анықталды. Демек, өсімдікте фотосинтез неғұрлым қарқынды өтетін болса, түсім солғұрлым жоғары болады.
Өсімдіктің ауа арқылы қоректенуі оның тамырмен қоректенуімен тығыз байланысты. Соның арқасында табиғатта ұдайы ауа алмасуы болып тұрады. Оны мына жағдаймен түсіндіруге болады. Жер бетінде жылына 400 млрд. тонна органикалық заттар түзіледі. Ол үшін өсімдіктер 170 млрд. тонна көмірқышқыл газын сіңіреді. Егер атмосферадағы көмірқышқыл газы жылма-жыл толықтырылмаса, оның қоры небәрі 4 жыл ішінде таусылған болар еді. Осыған орай өсімдіктерге қажетті көмірқышқыл газының қорын көбейту маңызды мәселе. Бұлардың негізгілері өсімдік пен жан-жануардың қалдықтарының шіріндісі болады. Мұнда атмосфераға көп мөлшерде көмірқышқыл газы бөлінеді. Топыраққа органикалық тыңайтқыш ендіру, жасыл тыңайтқышты тиімді қолдану, топырақ өңдеу жүйесін ұқыпты жүргізу, микробиологиялық процестің жедел жүруі де атмосферадағы көмірқышқыл газының қорының молаюына маңызды рөл атқарады.
3 Өсімдіктің қоректенуінің теориялары
Өсімдіктің тамыр арқылы қоректенуі .Жалпы өсімдіктің қоректенуі сыртқы факторға жатады. Өсімдіктің минералдық заттарды тамыры арқылы пайдаланылатыны жөнінде алғашқы деректер Дютроштың 1837 жылғы зерттеу жұмыстарында кездеседі. Ол өсімдік клеткаларының қоректік элементтерді қабылдауын диффузия құбылысымен, топырақтағы иондардың қозғалысын концентрациясы жоғары ерітіндіден концентрациясы төмен ерітіндіге жылжуымен түсіндірді.
Диффузиялық-осмостық теорияны жақтаушылар Де Фриз, Пфеффер, Майер өсімдіктің тамыры қоректік заттарды сумен бірге сорады деп түсіндіреді. Олардың пікірінше өсімдіктің қоректік заттарды сіңіруі транспирация құбылысына тікелей тәуелді. Шын мәнінде өсімдіктің қоректік заттарды қабылдауын тек қана бұл теориямен түсіндіру дұрыс емес. XIX ғасырдың аяғында Овертон өсімдіктің қоректенуі жөнінде өзінің липоидтық теориясын ұсынды. Бұл теория бойынша өсімдік клеткасына оның цитоплазмасының липидтік компонентінде еріген заттар енеді. Липоидтық теорияның диффузиялық-осмостық теориядан айырмашылығы еріген заттарды жекелеп пайдалануы, химиялық реакция арқылы цитоплазманың сыртқы қабатында жүреді деп түсіндіруінде.
Ультрафильтрация теориясын ашқан Траубе мен Руланд цитоплазма қабықшасы арқылы қоректік заттардың клеткаға енуі оның (цитоплазма қабықшасы) тесіктерінің көлемі мен сол заттардың молекулаларының мөлшеріне байланысты деп дәлелдеді. Өсімдіктің амин қышқылдарын, фитин және кейбір жоғары молекулалы органикалық заттарды сіңіруін бұл теориямен түсіндіру мүмкін емес. ХХ ғасырдың басында Дево өте сұйық ерітіндідегі катиондардың өсімдік клеткасымен жылдам байланысатынын анықтады. Бұл жағдай абсорбция теориясының пайда болуы мен дамуына әсерін тигізді. Дево өсімдік клеткасына енген катион эквивалентті мөлшерде оның құрамындағы басқа катиондарды ығыстратынын атап көрсетті. Бүл құбылыстың өзі ерітінді концентрациясы мен уақытқа байланысты өтеді.
1921-1928 жылдары Д. А. Сабинин клетканың заттарды бойына сіңіруі адсорбция реакциясы арқылы өтетіндігін анықтады. Д. А. Сабинин өсімдіктің қоректенуі жеке мүшелердің пайда болуы мен дамуының арасында байланыс бар екенін көрсетті. Д. А. Сабинин пікірі бойынша қоректік заттардың сіңірілуі олардың өсімдіктегі айналымының бір буыны болып табылады. Өсімдіктің заттарды сіңіруі оның жер бетіндегі мүшелерінің қызметіне, зат алмасудың қарқынына, ассимиляттардың тамырға жылжуына, тыныс алу процесінің деңгейіне байланысты. 1940 жылы Д. А. Сабинин тамыр арқылы өтетін заттардың өзгеруі туралы және синтетикалық қызметі жөнінде өз концепциясын ұсынды. Бүл концепцияның негізгі желісіне мыналар жатады:
1. Тамыр минералдық элементтерді сіңірумен қатар, оларды толық немесе жартылай басқа қосылыстарға айналдырып, өсімдіктің топырақ бетіндегі мүшелеріне ауыстырады.
2. Тамырдың синтетикалық қызметі фотосинтез процесімен байланысты.
3. Тамыр өсімдіктің жер бетіндегі мүшелеріне, оларды су және минералдық элементтермен қамтамасыз ету арқылы әсер етумен қатар зат алмасудың арнайы реакциясынан түзілетін фитогармондар арқылы да әсер етеді. Мүнда цитокининдер мен гибберлиндерді атауға болады.
Өсімдік тамыры қоректік заттарды сіңіретін басты мүше. Әрбір өсімдіктің тамыр жүйесінің құрылысы, топырақта таралуы бірдей емес. Өсімдік тамыры бірнеше қызмет атқарады. Атап айтқанда, сыртқы ортадағы су мен қоректік заттарды сіңіреді, онда көптеген биосинтетикалық процестер өтеді және тірек қызметін атқарады.
Тамырдың негізгі үш аймағы болады:
1. Өсу немесе бөліну аймағы. Ұзындығы 1,5 мм. Осы аймақта клеткалардың бөлінуі есебінен тамыр өседі;
2. Тамыр түкшелері немесе сору. Ұзындығы 1-2 см.
3. Созылу аймағы.
Бөліну немесе созылу аймақтарына қарағанда қоректік, элементтерді тамыр түкшелері орналасқан аймақ клеткалары әлдененше рет қарқынды сіңіреді. Өсімдіктің тамыр жүйелері құрғақ массаға шаққанда оның топырақ бетіндегі мүшелерінен үлкен болады. Мәселен, тың және тыңайған жерлерде өсірілетін көпжылдық шөптердің тамыр жүйелерінің үлес салмағы жер бетіндегі өсімдік массасының 90%-ін құрайды. Сол сияқты жүгері егістігінде ол 16%, күздік бидайда 70%, жоңышқада-166% жетеді. Ал жантақ өсімдігінің тамыр жүйесінің ұзындығы 30 метрге дейін жетіп, оның жалпы салмағы жер бетіндегі массасынан 100 есе артық болады. Ауыл шаруашылығы дақылдарының тамыр жүйелері біршама көлемді жерлерді алып жатады. Атап айтқанда, күздік қара бидай тамырларының ұзындығы 6,4 м, түкшелерінің ұзындығы 1649,4 метрге, олардың беткі қабатының көлемі 503-767 см2 жетеді. Бір күздік қара бидай өсімдігі 4 айда 13 млн 800 мың дана тамыр шығарады. Олардың жалпы ұзындығы 600 километрге, беткі жағының көлемі 240 шаршы метрге жетеді. Мақта тамыры 6 м, жаздық бидай тамыры 2 м, ал жоңышқа тамыры 18 м жетеді. Тамырдың ұзындығы көбінесе топырақтағы ылғал қорына байланысты болады. Мәселен, топырақта ылғал жеткілікті болған жағдайда жоңышқа тамыры 5 метр тереңдікке дейін, ал құаңшылық болған кезде 20 м тереңдікке дейін бойлап өседі. Табиғатта тамырлардың терең бойлап кетуіне жол бермейтін және оларды топырақтың жоғарғы бойына қабатында ғана өркен жаþға мәжбүр ететін факторлар болады. Бұған температурасы төмен және аграциясы нашар, ыза сулардың жер бетіне жақын орналасуын жатқызуға болады. Өсімдіктің тамыр түкшелері тамырдың қоректік заттарды сіңіру қабілетін ұлғайтады. Сондықтан да тамыр түкшелері орналасқан аймақты тамырдың қоректік заттарды сіңіру аймағы деп атайды. Дегенмен кейбір зерттеу жұмыстары қоректік заттарды сіңіруге тамыр түкшелері орналаспаған аймақ клеткалары да қатысатынын дәлелдеп отыр. Рассел мен Кларксон фосфор элементін арпа тамырының тамыр түкшелерінен 40 см қашығырақ орналасқан бөліктері де сіңіретінін анықтады. Сонымен қатар тамыр түкшелерінде арнайы сіңіру қасиеттері жоқ. Кларксон арпа өсімдігін тұздардың судағы ерітіндісінде өсіріп, ерітіндіні ұдайы араластырған. Арпа тамырында түкшелер пайда болмайды. Ал ерітіндідегі иондарды арпаның сіңіруі өте жоғары дәрежеде өтеді.
Дегенмен тамыр түкшелері қоректік заттарды жақсы сіңіреді және өздерінің өткізгіш жүйелері арқылы өсімдіктің жер бетіндегі мүшелерін қоректік заттармен қамтамасыз етеді. Ал созылу және бөліну аймақтарында өткізгіш жүйе жоқ. Бұл аймақтар сіңірген қоректік заттарды өздері пайданады.
Өсімдіктің қоректік заттарды сіңіруі жұмсалатын энергия түріне қарай активті және пассивті болып бөлінеді. Қоректік заттарды пассивті жолмен сіңіруге диффузия құбылысын жатқызуға болады. Диффузиялық қозғалыстың арқасында еріген заттардың молекулалары мен иондары бүкіл көлем бойына біркелкі таралады. Егер олардың концентрациясы көлемнің бір бөлігінде төмен‚ ал екінші бөлігінде жоғары болса‚ молекулалардың диффузиялық қозғалысы иондардың концентрациясы аз бағытта басым болады. Сөйтіп бұл қозғалыс заттардың әртүрлі бөліктеріндегі концентрациясы теңескенше жалғаса береді. Жүйенің жеке бөліктеріндегі заттардың концентрация айырмашылығы концентрация градиенті деп аталады. Бұл жағдайда уақыт бірлігі ішінде, заттардың молекуласы мен иондары ерітіндінің концентрациясы төмен бағытына қарай қозғалады да, концентрация градиенті теңеседі. Концентрацияның бұл градиентін химиялық потенциал градиенті деп атайды.
Диффузия жылдамдығы градиенттің шамасына‚ молекуланың мөлшері мен массасына‚ олардың химиялық табиғатына‚ ортаның тұтқырлығына‚ температураға‚ электростатистикалық күштерге байланысты. Зарядталған иондар диффузияланған кезде, олардың градиенті жүйеде электрохимиялық градиент туғызады. Электрохимиялық градиент мынадай жағдайда пайда болады: бір заттың анионы мен катионының диффузиясы бір-біріне әсер етуі мүмкін және олардың диффузиясының жылдамдығы түрліше болуы ықтимал. Осының салдарынан жүйеде электр зарядтары біркелкі таралмайды.
Сонымен клеткалардағы молекулалар мен иондардың қозғалуы химиялық потенциал градиенті мен электрлік потенциал градиентіне байланысты. Иондарды тасымалдайтын бұл күшті электрохимиялық қозғаушы күш дейді. Егер диффузия мембрана арқылы жүретін болса және анион мен катион мембранадан түрліше жылдамдықпен қозғалса, мембрана бетінде түрлі мөлшерде заряд жиналып, мембраналық электр потенциалын туғызады.
Сіңіру процесі заттардың клеткаға активті жолмен тасымалдануы арқылы да жүзеге асады. Активті тасымалдау клетканың тіршілік жағдайларына байланысты.
Енді иондардың тамыр клеткасына қарай активті жолмен өтуінің кейбір ерекшеліктеріне тоқталайық. Плазмалеммадан тыс орналасқан клетка аралықтары және клетка қабықшасындағы фибрилла мен мицелла аралықтары біртұтас бос кеңістік түзеді. Оны апопласт деп атайды. Барлық клеткалардың апопластары өзара байланысып біртұтас жүйе құрайды. Сыртқы ортадағы иондар апопластқа диффузия немесе гидростатикалық қысым әсерінен қозғалатын су ағынымен бірге жылжиды. Апопластан өтетін су иондарды өзімен бірге әкетеді. Клетка қабықшасы теріс зарядты болғандықтан көптеген катиондарды адсорбциялайды‚ сөйтіп олардың концентрациясын 20-800 есе жоғарлатады. Иондар концентрациясының көтерілуі олардың одан ары жылжуын жеңілдетеді. Қабықшаға енген иондар апопласт жолына түседі немесе плазмалемма арқылы өтіп симпласт жолына енеді. Сонымен қатар ерітінді қабықшаның тесіктері арқылы өтіп‚ плазмалеммамен тікелей жанасуы да мүмкін. Мұнда да иондар симпласт жолына өтеді. Симпласт деп өзара жалғасып жатқан цитоплазманың плазмадесма жіпшелерімен байланысуынан пайда болған торды айтады. Симпласт арқылы тасымалданған иондар метаболизмге қосылады және шоғырлана келе концентрациясы артады. Иондардың мембрана арқылы екі рет өтіп‚ тамырдың түкшелеріне шығуы екі түрлі насос күшімен жүреді. Бірінші насос тамырдың эпидермис клеткаларының плазмолеммасында орналасқан. Ол иондарды сыртқы ортадан тамыр клеткасына тасымалдайды. Екінші насос ксилемаға жанасып жатқан клеткалардың плазмолеммасында орналасады. Бұл насос иондарды ксилема түктеріне шығарады.
Өсімдік қоректік элементтерді тамырдың топырақпен және топырақ ерітіндісімен тікелей жанасуы арқылы сіңіреді. Қоректік зат тамыр арқылы адсорбциялық жолмен енеді. Тамыр клеткаларындағы белок құрамында негіздік және қышқылдық топтардың болуынан протоплазма бетінде оң және теріс зарядты учаскелер болады. Демек протоплазма қоректік заттан бір мезгілде түрлі катиондар мен аниондарды адсорбциялайды. Мұнда өсімдік тамыры тыныс алған кезде пайда болған көмірқышқыл газы өсімдік тканьдарындағы сумен әрекеттеседі де көмір қышқылы (Н2СО3) түзіледі. Бұл қосылыс диссоцациялануынан Н+ ионы топырақтағы катиондармен (K+, Ca2+ , Mg2+ ,NH4-), HCO3- аниондармен (NO-3 ‚ SO42-‚ PO3-4) алмасады. Топырақ ерітіндісінен тамырға өткен иондар ішінара басқа қосылыстарға айналады немесе өзгеріссіз сабақтар мен жапырақтарға тарайды.
Сонымен қатар‚ тамырдың өзі органикалық қышқылдар бөліп‚ олар да шамалы еритін қосылыстарды сіңіруге ықпал етеді.
Н+ және НСО-3 иондарының сіңірілуі жанасу арқылы да болады. Олар топырақтың қатты фазасында болатын иондармен тікелей алмаса алады.
Кез келген өсімдік қоректік элементтерді пайдаланғанда өзінің биологиялық ерекшеліктеріне қарай талғампаздық танытады.