Фізіологія збудження
Як було вказано раніше, подразливість — одна із основних властивостей кожної клітини організму. Завдяки цій властивості організм реагує на дію зовнішнього чи внутрішнього середовища збудженням, тобто переходить із стану фізіологічного спокою в діяльний стан. Процеси, що протікають у клітинах при збудженні, характеризуються сукупністю хімічних, структурних, температурних, функціональних та електричних змін.
Електричні зміни, або електричні імпульси, є однією з найбільш .характерних ознак збудження, вони мають ще назву біоелектричних явищ. Біоелектричні явища, що спостерігаються в нервових структурах, одержали назву нервових імпульсів, завдяки яким нервові клітини включають в дію ефектори — м'язи, залози. Нер-вові імпульси є тією «мовою мозку», на якій він виконує всі свої функції — сприйняття інформації, аналіз її, зберігання і синтез.
Здатність клітин сприймати подразнення і переходити із стану спокою в діяльний стан зумовлюється будовою і властивостями клітинної мембрани.
Структура і властивості клітинних мембран.Клітини кожної тканини організму оточені осмотично-активною мембраною, або плазматичною мембраною. Під електронним мікроскопом вона має простий вигляд. Загальна її товщина 7,5 ... 18 нм. У відповідності з сучасними уявленнями, мембрана складається з трьох шарів (рис. 6): біомолекулярного шару ліпідів, який розташований між
21 
двома шарами білків. Мембрана має отвори, канали діаметром 0,7 ... 0,8 нм. Існує думка, що ці канали несуть електричний заряд і що саме це дозволяє вибірково пропускати одні іони в клітину і затримувати інші.
З функціональної точки зору мембрана не тільки перетинка, значення якої обмежується механічною функцією — наданням " певної форми цитоплазмі. Вона є надзвичайно важливим утворенням живої клітини і виконує багато функцій. Клітинна мембрана вибірково регулює надходження молекул та іонів в клітину і з неї, їх кількісний і якісний склад, тобто «вирішує», які саме молекули необхідно пропустити всередину клітин, а які необхідно залишити за її межами, які молекули необхідно залишити в клітині, а які видалити з неї.
Крім зовнішньої плазматичної мембрани є внутрішні мембрани, які розподіляють клітину на «засіки». Це дозволяє накопичувати в окремих ділянках клітини одні речовини і не впускати туди інші. Таким чином, завдяки мембранам виникає морфологічна основа для внутрішнього клітинного розподілу біохімічних реакцій. Цитоплазматичні мембрани виконують функцію насоса, який накачує в клітину або викачує з неї фізіологічно активні іони. Клітинна мембрана має особливі каталітично активні ділянки, а також механізми, які за рахунок енергії АТФ виконують осмотичну роботу проти градієнта концентрації. На мембранах мітохондрій і інших органоїдів проходять процеси перетворення енергії і біосинтезу. Нарешті, мембрана — надчутливий сприймач і перетворювач світлових, звукових, механічних, хімічних сигналів зовнішнього світу в специфічну активність клітин — електричні явища.
Біоелектричні явища. Мембранний потенціал. Після відкриття біоелектричних явищ стало зрозумілим, що виникнення і розповсюдження збудження в збудливих тканинах пов'язано саме з ними. Тому розкриттю механізмів їх виникнення було присвячено безліч дослідів починаючи з кінця минулого століття і протягом всього теперішнього століття. Але тільки завдяки запровадженню мікроелектродної техніки в останні двадцять років стало можли-
22 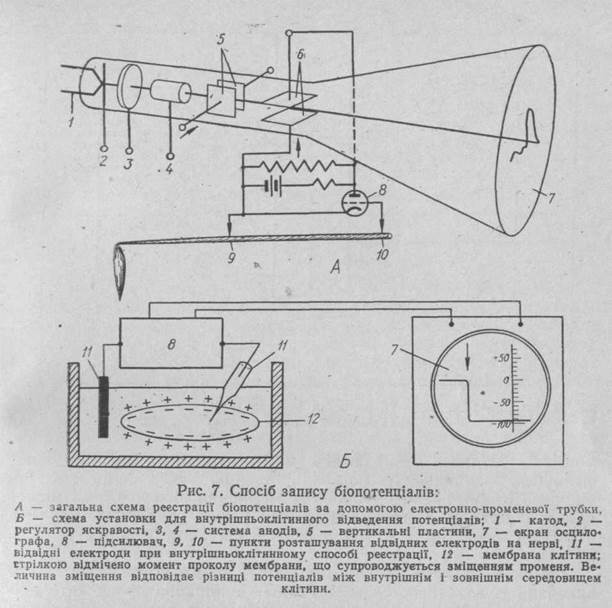
вим перейти від гіпотетичного уявлення механізмів до твердого розуміння цього явища. Суть цієї техніки полягає в тому, що в середину клітини вводиться дуже тонкий, 0,5 мкм в діаметрі, електрод, за допомогою якого можна відводити електричні потенціали тільки однієї клітини без суттєвих перешкод з боку інших клітин (рис. 7). Реєстрування потенціалів здійснюється за допомогою катодного осцилографа. Мікроелектродна техніка дозволяє також наносити подразнення в певних частинах клітини і водночас відходити на реєструючі прилади відповіді на подразнення. А саме це дає змогу вивчати функціональні властивості клітини при їх діяльності в бажаному для експериментатора напрямку.
За допомогою введення мікроелектродів безпосередньо в клітину встановлено, що внутрішня сторона мембрани нервової або
23 
м'язової клітини в стані спокою має негативний заряд по відношенню до зовнішньої її поверхні. Різницю потенціалів між зов-.. нішньою і внутрішньою сторонами мембрани називають мембранним потенціалом. Його величина для різних збудливих структур лежить в межах 60 ... 90 мВ, а для епітеліальної тканини — 18 ... 20 мВ.
Виникнення мембранного потенціалу спокою можна пояснити виходячи із властивостей мембрани — вибірково пропускати або затримувати різні іони. Клітинні мембрани в спокої дрбре проникні для іонів К+ і мало проникні для іонів Na+ і аніонів (А-) органічних речовин. Внаслідок процесів активного транспорту іонів через мембрану "концентрація іонів К+ в середині клітини в 30... 50 разів вища, ніж у позаклітинній рідині. Концентрація Na+в 10 разів, а СГ- в 50 разів більша зовні клітини. Переважна більшість аніонів органічних молекул знаходиться в середині клітин (рис. 8). Така асиметрія концентрацій різних іонів в середині ї вовні клітини є передумовою виникнення мембранного потенціалу.
Іони К+, внаслідок їх легкої проникності через мембрану, постійно виходять із клітин, де концентрація їх висока, в позаклітинний простір. Виносячи позитивний заряд назовні, іони К-збільшують мембранний потенціал, іони Na+, хоч і мають меншу проникність, ніж іони К+, все ж надходять в середину клітин і

зменшують його. Таким чином, величина мебранного потенціалу клітини зумовлюється швидкостями проходження через мембрану іонів Na+ і К+. Чим більше іонів К+ за одиницю часу виходить назовні і менше входить всередину клітин іонів Na+, тим більший мембранний потенціал. Для підтримання різниці концентрації іонів, а значить, і різниці потенціалів на мембрані, мають значення не тільки пасивні дифузійні механізми, а також і активні механізми, які здатні повертати іони К+ в клітину проти градієнта його концентрації. Такі ж механізми існують і для активного переносу Na+ із середини клітини на поверхню її мембрани. Якби не було цих механізмів, то уже через одну годину кількість іонів Na+ в середині клітини збільшилась би вдвічі. Про це свідчать спеціальні розрахунки. В дійсності такого збільшення не відбувається завдяки підтриманню градієнта концентрацій Na+ і К+ спеціальними механізмами активного їх переносу через мембрану. Ці механізми називають натрієвою і калієвою помпами.
Активне переміщення іонів здійснюється за рахунок енергії АТФ. При отруєнні збудливої тканини ядами, що порушують обмін речовин, наприклад цианідами, дінітрофенолом, активний транспорт іонів припиняється і мембранний потенціал зменшується. При введенні в клітину АТФ іони Na+ знову починають повертатися із середини клітини в оточуюче середовище, градієнт його концентрації відновлюється.
Потенціал дії.При подразненні нервових або м'язових клітин мембранний потенціал їх зменшується, оскільки мембрана їх де-поляризується, при цьому виникає хвиля збудження. Короткочасна зміна мембранного потенціалу носить назву потенціалу дії, або електричного імпульсу. Потенціал дії має характерну структуру (рис. 9). Розрізняють короткочасний пік, який складається із висхідної і низхідної частин, і слідові потенціали — позитивний і
ч 
негативний. В основі виникнення потенціалу дії лежить зміна проникності мембрани для іонів Na+ і К+. Під час подразнення клітини проникність Na+ через мембрану збільшується у 20 разів, а вихід К+ назовні тільки в 9 разів. Цей факт має велике значення для виникнення потенціалу дії. Іони Na+, проникаючи в клітину через мембрану, нейтралізують негативний внутрішній заряд, а це, в свою чергу, викликає ще більше, лавиноподібне проникнення їх в клітину і деполяризацію мембрани.
При цьому іонів Na+ надходить така кількість, що настає інверсія (зміна знака) мембранного потенціалу. Зовнішня частина мембрани приймає негативний, а внутрішня позитивний знак. Перехід іонів Na+ всередину клітини викликає електричний струм, направлений всередину, і зумовлює висхідну фазу потенціалу дії, а вихід іонів К+ забезпечує реполяризацію мембрани і зумовлює низхідну фазу потенціалу дії. Пік потенціалу дії залежно від функціональних властивостей тканин коливається в межах 100... 120 мВ, а його тривалість —0,5 ... 5 мс. Після закінчення піку потенціалу дії мембрана протягом 15... 30с залишається ще частково деполяризованою, що позначається як слідовий потенціал. Його виникнення пов'язано з тим, що вхід іонів Na+ в клітину в цей час зменшується, а вихід іонів К+ збільшується (рис. 10). А оскільки струми Na+ і К+ протилежно направлені, то співвідношення їх підтримує мембрану частково деполяризованою. Поступово надходження іонів N+ в середину клітини припиняється зовсім, а іони К+ продовжують ще виходити назовні, виносячи позитивний заряд, тим самим збільшуючи мембранний потенціал вище того рівня, який був до збудження. Цей період називається періодом гіперполяризації мембрани, або слідовим позитивним потенціалом. Його тривалість може бути різною, від 50 до 300 мс, залежно від функціональних властивостей нервових клітин. По закінченні потенціалу дії включається натрієвий насос (помпа), який викачує іони назовні і тим самим відновлює вихідну різницю концентрації. Робота натрієвого насоса забезпечується за рахунок енергії АТФ. Кількість енергії, яка витрачається на хімічні і електричні процеси, пов'язана з виникненням і проведенням нервових імпульсів і дуже мала. Нервове
волокно, в якому виключені механізми утворення АТФ, може провести більше 100 тис. імпульсів.
Збудливість і її параметри. Встановлено, що потенціал дії виникає тільки тоді, коли подразник досягає певної сили і діє певний - час. Мінімальна сила подразника, яка необхдіна, щоб викликати збудження тканин, характеризує її рівень збудливості і називається пороговою силою. Але навіть при дії підпорогових подразників на мембрані відбуваються значні зміни, що передують появі потенціалу дії. Це так звані місцеві потенціали мембрани, або локальні відповіді. Величина їх залежить від сили подразника. Вони здатні до сумації.
Як тільки місцевий потенціал досягає певної величини (для нервових волокон приблизно 10 мВ), виникає потенціал дії. Величину місцевого потенціалу, яка призводить до появи потенціалу дії, називають пороговим потенціалом, а рівень деполяризації мембрани, що досягається при цьому, — критичним рівнем деполяризації (рис. 11).
Важливе значення для виникнення збудження в м'язах і нервах має також швидкість зростання сили струму, тобто його градієнт. При дуже повільному зростанні сили струму потенціал дії не виникає. Пристосування збудливих тканин до дії повільно зростаючого подразника дістало назву акомодації (рис. 12). Це явище пов'язане з тим, що збільшення проникності іонів Na+ за час дії подразника компенсується виходом іонів К+, пороговий потенціал підвищується і не досягається критичний рівень деполяризації мембрани.
Механізм розповсюдження збудження. Однією з характерних рис збудливих тканин є розповсюдження збудження. Потенціал дії, який


27 
виникає при збудженні, здатний до розповсюдження за рахунок тих електричних струмів, які він викликає.
В стані спокою зовнішня сторона мембрани заряджена позитивно, а внутрішня — негативно. Під час збудження знак заряду на мембрані змінюється на протилежний, тобто зовнішня сторона мембрани стає негативно зарядженою, а внутрішня — позитивно. Внаслідок цього між збудженою і незбудженою ділянками вздовж мембрани виникає різниця потенціалів, яка призводить по переміщення іонів (рис. 13). На поверхні мембрани позитивні іони рухаються від незбудженої ділянки до збудженої, а на внутрішній стороні мембрани, навпаки, позитивні іони рухаються від місця збудження до незбудженої ділянки. Рух іонів утворює колові електричні струми, які деполяризують мембрану на незбудженій ділянці. Коли деполяризація досягне критичного рівня, виникає новий регенеративний потенціал дії і процес збудження переміститься на наступну ділянку. Далі виникає ланцюгова реакція. Потенціал дії, «самовідтворюючись», розповсюджується по нервовому або м'язовому волокні. Швидкість розповсюдження нервового імпульсу залежить від функціональних властивостей і коливається в межах 2 ... 120м/с.
Зміни збудливості тканин в процесі розвитку хвилі збудження. Впроцесі розвитку хвилі збудження в збудливих тканинах виникають закономірні зміни їхньої збудливості. Про це свідчать досліди з подразненням нервових волокон короткими імпульсами, які йдуть один за другим з певними проміжками часу.
В період дії подразника, коли на мембрані розвиваються локальні зміни мембранного потенціалу дії, збудливість підвищена (рис. 14). Під час розвитку піку потенціалу дії збудливість мембрани падає до нуля, тобто в цей час найсильніший подразник не здатний викликати нового збудження. Цей період називається фазою абсолютної незбудливості, або абсолютної рефрактерності. Тривалість її дорівнює тривалості піку потенціалу дії (для волокон рухових нервів — 0,5 мс). Фаза абсолютної рефрактерності змінюється фазою відносної рефрактерності. Збудливість в цій фазі поступово відновлюється,

Рис. 14. Зміна збудження при проходженні хвилі збудження, А — компоненти хвилі збудження: 1 — деполяризація, 2 — реполяризація, МК — критичний рівень деполяризації, а — тривалість порогового потенціалу, б — тривалість потенціалу дії. в — слідова негативність, г — слідова позитивність; Б — зміни збудливості в різні фази хвилі збудження: Р3 — рівень збудливості в спокої, а — підвищення збудливості до нуля в період порогового потенціалу, б — падіння збудливості до нуля в період протікання потенціалу дії (абсолютна реф-рактерність); в1 — повернення збудливості до вихідного рівня в період слідової негативності (відносна рефрактерність), в2 — підвищення збудливості на кінець слідової негативності (екзальтація, або супернормальність), в — весь період слідової негативності, г — падіння збудливості в період гіперполяриза-ції (субнормальність).
але для виникнення потенціалу дії сила подразника повинна бути більшою від порогової. Тривалість цієї фази 4 ... 8 мс. Після фази відносної рефрактерності збудливість тканини на деякий час стає більшою від вихідної. Цей період дістав назву супернормаль-ної фази. В її основі лежить слідова деполяризація мембрани. В швидких рухових нервових волокнах людини ця фаза триває 15... 30 мс. Підвищена збудливість (супернормальна фаза) змінюється невеликим зниженням збудливості — субнормальною фазою, виникнення якої пов'язане з гіперполяризацією мембрани. Тривалість її в рухових нервах 50 мс. По закінченні цієї фази встановлюється початкова збудливість тканин.
Функціональна рухливість (лабільність). В природних умовах у збудливих тканинах спостерігаються не поодинокі імпульси, а серія їх з певним інтервалом між імпульсами. Залежно від властивостей і функціонального стану збудливі тканини можуть сприймати лише певну частину імпульсів. Тому М. Є. Введенським для характеристики швидкості протікання процесів збудження в тканинах було запропоновано поняття функціональної рухливості, або лабільності. Функціональна рухливість вимірюється тією максимальною кількістю потенціалів дії, яку збудлива тканина може відтворити за одну секунду у відповідності з частотою її подразнення. Чим коротша рефрактерна фаза збудливої структури, тим вища її лабільність. Найвища лабільність у рухових нервових волокон теплокровних — 1000 імпульсів у секунду, волокон симпатичної нервової системи — 500 імпульсів у секунду, м'язових волокон — 200, а нервово-м'язових синапсів — лише 100 імпульсів у секунду.
Величина лабільності збудливих структур може збільшуватись або зменшуватись. При виконанні фізичних вправ швидкісного характеру лабільність м'язів підвищується. Підвищення лабільності може привести до того, що нервові клітини або м'язи почнуть відповідати на більш високу частоту подразнень. Це явище називають засвоєнням ритму (О. О. Ухтомський). Тривала робота викликає зниження лабільності нервових центрів і м'язів внаслідок розвитку в них втоми.