Митотический цикл.Характеристика интерфазы
Интерфаза
(от лат. inter — между и Фаза)
интеркинез, стадия жизненного цикла клетки между двумя последовательными митотическимиделениями (см. Митоз). Обычно различают гетеросинтетическую И., когда клетка растет, дифференцируется,осуществляет свойственные ей функции, и автосинтетическую И., в течение которой происходит подготовкаклетки к следующему делению. В зависимости от интенсивности синтеза дезоксирибонуклеиновой кислоты(См. Дезоксирибонуклеиновая кислота) (ДНК) автосинтетическую И., в свою очередь, делят на 3 периода: G1— предсинтетический, или постмитотический, S — синтетический и G2 — предмитотический, илипостсинтетический. В G1-периоде осуществляются накопление необходимых для деления клеткиэнергетических ресурсов, синтез рибонуклеиновой кислоты (См. Рибонуклеиновые кислоты) (РНК), идётподготовка к удвоению молекул (репликации (См. Репликация)) ДНК; за счёт синтезированного в этот периодбелка увеличивается масса клетки и образуется ряд ферментов, необходимых для синтеза ДНК вследующем периоде И. В S-периоде происходит синтез ДНК, т. е. осуществляется репликация её молекул. ВG2-периоде синтез ДНК закончен, усиливается синтез РНК и белков, видимо, идущих на построениемитотического аппарата (См. Митотический аппарат).
В клетках взрослого организма И, продолжается от 10 до 30 часов и больше; в быстроделящихсяклетках И. длится несколько минут (например, в яйцах морского ежа на стадиях 2—4 бластомеров — 14мин.).
Некоторые авторы выделяют в И. «нулевой период» — G0 предшествующий периоду G1. Наиболееотчётливо G0 проявляется в клетках, которые во взрослом организме, как правило, не делятся (под влияниемразных факторов они могут войти в G1-период, пройти затем периоды S, G1 и вступить в собственно митоз).Резкой границы между всеми периодами И., а также между митозом и И. не существует. Ряд авторовполагает, что периоды S и G2 следует считать началом митоза — препрофазой, с которой и начинаетсярепродукция клетки, а не относить их к И.
МИТОЗ
(от греч. mitos — нить), непрямое деление, основной способ деления эукариотных клеток. Биол. значение М.состоит в строго одинаковом распределении редуплицированных хромосом между дочерними клетками, чтообеспечивает образование генетически равноценных клеток и сохраняет преемственность в ряду клеточныхпоколений. В 1874 И. Д. Чистяков описал ряд стадий (фаз) М. в спорах плаунов, ещё не ясно представляясебе их последовательность. Детальные исследования по морфологии М. впервые были выполнены Э.Страсбургером на растениях (1876—79) и В. Флеммингом на животных (1882). Продолжительность М. всреднем 1—2 ч. В процессе М. условно выделяют неск. стадий, постепенно и непрерывно переходящих другв друга: профазу, прометафазу, метафазу, анафазу и телофазу. Длительность стадий М. различна и зависитот типа ткани, физиол. состояния организма, внеш. факторов; наиб, продолжительны первая и последняя.Важнейшие признаки профазы — конденсация хромосом, распад ядрышек и начало формированияверетена деления, снижение активности транскрипции (к концу профазы синтез РНК прекращается).Веретено деления образуется либо с участием пентриолей, образуя митотический аппарат (в клеткахживотных и нек-рых низших растений), либо без них (в клетках высших растений и нек-рых простейших). Уводорослей, низших грибов и ряда простейших веретено может формироваться внутри ядра (т. п. закрытыйМ.). Прометафаза начинается распадом ядерной оболочки на фрагменты и беспорядочными движениямихромосом в центр, части клетки, соответствующей зоне бывшего ядра. При «закрытом М.» оболочка ядрасохраняется в течение всего М. в метафазе завершается формирование веретена деления. Хромосомыперестают двигаться и выстраиваются по экватору веретена, образуя экваториальную пластинку. Синтезбелка снижен на 20—30% по сравнению с интерфазой. На этой стадии М. клетки наиб, чувствительны кхолоду, колхицину, его производным и др. агентам, воздействие к-рых разрушает веретено деления иприводит к прекращению деления клеток (К-митоз). При низких дозах повреждающих агентов нормальноетечение М. восстанавливается через неск. часов после их воздействия; более высокие дозы приводят либо кгибели клетки, либо к её полиплоидизации.Анафаза — самая короткая стадия М. Характеризуетсяразделением сестринских хроматид и расхождением хромосом к противоположным полюсам клетки.Скорость их движения в среднем 0,2—5 мкм/мин. В ряде случаев движение хромосом к полюсам клеткисопровождается дополнит, расхождением полюсов друг от друга. Телофаза длится с момента прекращениядвижения хромосом до окончания процессов, связанных с реконструкцией дочерних ядер (десприрализацияи активизация хромосом, образование ядерной оболочки, формирование ядрышек), с разрушениемверетена деления, разделением тела материнской клетки на 2 дочерние и образованием (в клеткахживотных) остаточного тельца флемминга. По завершении цитотомии клетки вступают в интерфазу, к-раяначинается G1-периодом следующего клеточного цикла. В опытах с температурно-зависимыми мутантамидрожжей и клеточных линий млекопитающих показано, что протекание М. обусловливается активациейопределённых генов и синтезом специфич. РНК и белка. Иногда М. считают только деление ядра(кариокинез), к-рое не всегда сопровождается цитотомией — образованием двух отд. клеток. (см. МЕЙОЗ,ЭНДОМИТОЗ, АМИТОЗ).
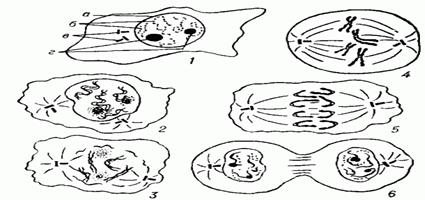
Общая схема митоза: 1 — интерфаза; 2 — профаза; 3 — прометафаза; 4 — метафаза; 5 — анафаза; 6 —телофаза; а — ядерная оболочка; б — хромосомы; в — центриоли; г — ядрышки.
.(Источник: «Биологический энциклопедический словарь.» Гл. ред. М. С. Гиляров; Редкол.: А. А.Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Сов. Энциклопедия, 1986.)
мито́з
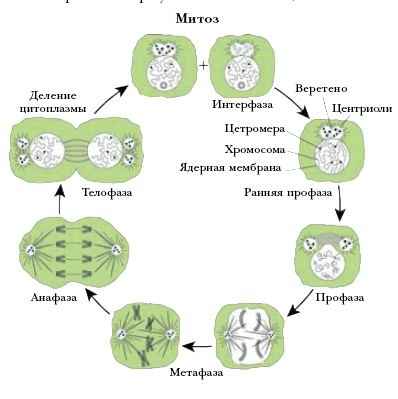
способ деления клеток, при котором генетический материал (хромосомы) распределяется поровну междуновыми (дочерними) клетками. Начинается с разделения ядра на два дочерних. Аналогично делится ицитоплазма. Процессы, происходящие от одного деления до другого, называются митотическим циклом. Онсостоит из 2 стадий – интерфазы (стадии покоя) и собственно митоза (стадии деления). В интерфазе в клеткепроисходит образование ДНК. Интерфаза делится на 3 периода. В первый период, продолжающийся 12—24ч, происходит накопление РНК и белков. Второй период (синтетический) характеризуется образованием ДНК,в результате чего её количество удваивается. В течение третьего периода (постсинтетического) происходитнакопление энергии, после чего клетка из стадии интерфазы переходит к митозу. Митоз проходит 4последовательные фазы – профазу, метафазу, анафазу и телофазу. В профазе хромосомы уплотняются,скручиваются в спирали и становятся видимыми под микроскопом. Мембрана ядра растворяется поддействием ферментов, ядрышко исчезает. Центриоли начинают расходиться к полюсам. Между полюсамиформируется веретено деления клетки – структура, состоящая из РНК и белка. К концу профазы хромосомыудваиваются, но члены каждой пары удерживаются рядом. В метафазе они располагаются по экваторуклетки. Хроматиды прикреплены к нитям веретена и уже начинают отсоединяться. В анафазе каждаяхроматида приобретает собственную центромеру, удлиняется и становится дочерней хромосомой. Нитиверетена, прикреплённые к центромерам, разводят «молодые» хромосомы к полюсам клетки. В телофазедочерние хромосомы достигают полюсов, их спирали раскручиваются, удлиняются и опять становятся плоховидимыми в микроскоп. Образуется ядерная оболочка, вновь появляется ядрышко. В результате клеткаимеет двойное количество клеточных структур и общую цитоплазму. В конце митоза происходит её деление.В экваториальной зоне клетки образуется перетяжка, делящая её на 2 дочерние. У растений на местеперетяжки образуется пластинка из целлюлозы.
Продолжительность митотического цикла у разных клеток различна (от нескольких часов до нескольких дней)и зависит от многих факторов: температуры, физиологиче-ского состояния организма и др. Разные тканиобладают разной митотической активностью. В стабильных (мышцы, нервная система) клетки не делятся, алишь подвергаются возрастным изменениям (стареют). Растущие ткани содержат клетки, не обладающиемитотической активностью, и клетки, делящиеся посредством митоза. В результате органы из этих тканейспособны к росту. Обновляющиеся ткани (кожи, костного мозга, кишечника) содержат клетки, постоянноделящиеся в течение всей жизни организма.
74.
 Расшифровка регуляции процессов клеточного деления началась в 70е годы прошлого века, когда были найдены методы слияния разных клеток, методы получения гетерокарионов (о них см. главу 2). Оказалось, что можно получить слияние не только интерфазных клеток с интерфазными, но и интерфазных с митотическими клетками (рис. 114). При этом обнаружилось удивительное удивительное явление: в такой гибридной клетке, где были митотические хромосомы одной из исходных клеток, в ядрах от интерфазных клеток начиналась конденсация хромосом, разрушалась ядерная оболочка и образовывались т.н. преждевременно конденсированные хромосомы (ПКХ или РСС – preliminary condenced chromosomes). Причем структура ПКХ зависела от стадии интерфазного ядра: в ядре от G1 клетки появлялись однонитчатые длинные ПКХ, в ядре от G2 клетки ПКХ были двойными (т.к. после S-периода хромосомы удвоились). Эти наблюдения наводили на мысль, что при слиянии таких клеток на интерфазные ядра действуют какие-то факторы, содержащиеся в митотических клетках.
Расшифровка регуляции процессов клеточного деления началась в 70е годы прошлого века, когда были найдены методы слияния разных клеток, методы получения гетерокарионов (о них см. главу 2). Оказалось, что можно получить слияние не только интерфазных клеток с интерфазными, но и интерфазных с митотическими клетками (рис. 114). При этом обнаружилось удивительное удивительное явление: в такой гибридной клетке, где были митотические хромосомы одной из исходных клеток, в ядрах от интерфазных клеток начиналась конденсация хромосом, разрушалась ядерная оболочка и образовывались т.н. преждевременно конденсированные хромосомы (ПКХ или РСС – preliminary condenced chromosomes). Причем структура ПКХ зависела от стадии интерфазного ядра: в ядре от G1 клетки появлялись однонитчатые длинные ПКХ, в ядре от G2 клетки ПКХ были двойными (т.к. после S-периода хромосомы удвоились). Эти наблюдения наводили на мысль, что при слиянии таких клеток на интерфазные ядра действуют какие-то факторы, содержащиеся в митотических клетках.
Для проверки этого предположения были поставлены опыты со слиянием клеток на разных стадиях клеточного цикла (см.табл.)
Таблица. Результаты слияния клеток на разных стадиях (G1, S, G2 ,M) клеточного цикла
| Исходные клетки | гетерокарион |
| G2® G1 | Нет влияния |
| S ® G1 | Синтез ДНК |
| S ® G2 | Нет влияния |
| M ® G1 | М (ПКХ) |
| M ® S | М (ПКХ) |
| M ® G2 | М (ПКХ) |
Эти эксперименты были дополнены тем, что часть цитоплазмы митозной клетки инъецировали в цитоплазму G1-интерфазной клетки. Это приводило к появлению в ядре интерфазной клетки ПКХ. Из этих наблюдений был сделан ввод о том, что в цитоплазме митотической клетки есть фактор (или факторы), стимулирующие митоз (ФСМ или MPF – mitosis promoting factor). Этот фактор вызывает не только конденсацию хромосом, но и приводит к распаду ядерной оболочки, т.е. переводит интерфазную клетку, даже без синтеза ДНК, в митотическое состояние (конечно, дальше появления конденсированных хромосом развитие митоза не идет).
Параллельно с этими наблюдениями были проведены эксперименты на созревающих и дробящихся яйцеклетках лягушек X.laevis (рис. 346).
Ооцит X.laevis после репликации ДНК и короткой G2–стадии переходит в мейотическую профазу I, во время которой в течение 8 месяцев происходит рост и созревание ооцита, он дорастает до размера зрелой икринки. При спаривании овариальные клетки самки выделяют гомон прогестерон, который стимулирует переход из профазы в I мейотическое деление (митоз I), затем после короткой интерфазы наступает II мейотическое деление, которое на некоторое время останавливается на стадии метафазы (стадия яйца). При оплодотворении спермием метафаза завершается, происходит второе деление меойза, после чего гаплоидное ядро яйцеклетки сливается с ядром спермия, и образуется диплоидная зигота. После этих событий следуют через каждые 30 мин многочисленные деления клеток развивающейся бластулы (рис. 346а).
Если взять с помощью микроманипулятора небольшую часть цитоплазмы из ооцита на стадии метафазы II мейотического деления и инъецировать ее в цитоплазму не стимулированного прогестероном ооцита, то произойдет повторение описанного выше процесса: ооцит вступит в I деление мейоза, а затем и во II деление, т.е. произойдет его созревание (рис. 346в). Таким образом было найдено, что в ооците на стадии метафазы II деления в цитоплазме существует фактор (или факторы), стимулирующие созреваниеяйцеклетки (ФСС или MPF – maturation promoting factor).
Оказалось, что этот фактор (будем называть его MPF) присутствует в клетках только во время митотического состояния. Он обнаруживается также и во время дробления яйцеклетки (рис. 347). Таким образом, уровень MPF в интерфазных клетках низкий, а в митотических высокий.
Далее было найдено, что при инъекции цитоплазмы из митотических клеток культуры ткани в нестимулированный ооцит X. laevis, происходит созревание ооцита. Следовательно, фактор, стимулирующий митоз и фактор, стимулирующий созревание ооцитов – одно и тоже.
Этот фактор, MPF, был выделен и охарактеризован. Это гетеродимерный комплекс, состоящий из белка циклина (см. ниже) и зависимой от циклина протеинкиназы (Cyclin dependent kinase – Сdk), фермента, относящегося к фосфорилазам, который модифицирует белки, перенося фосфатную группу от АТФ на аминокислоты серин и треонин. Следовательно, MPF состоит из двух субъединиц: каталитической (Сdk) и регуляторной (циклин) (рис. 348).
Клеточный цикл, периоды
Клеточный цикл включает строго детерминированный ряд последовательных процессов, согласно позиции Hartwellа, 1995. Клетка должна между двумя последовательными делениями удвоить все свои компоненты и свою массу. Таким образом клеточный цикл составляют два периода:
1) период клеточного роста, называемый " интерфаза ", и
2) период клеточного деления, называемый " фаза М " (от слова mitosis). В свою очередь, в каждом периоде выделяют несколько фаз (рис.3).
Обычно интерфаза занимает не меньше 90% времени всего клеточного цикла. Например, у быстро делящихся клеток высших эукариот последовательные деления происходят один раз в 16-24 часа, и каждая фаза М длится 1-2 часа. Большая часть компонентов клетки синтезируется на протяжении всей интерфазы, это затрудняет выделение в ней отдельных стадий по мнению Pardee, 1989. В интерфазе выделяют фазу G1, фазу S и фазу G2. Период интерфазы, когда происходит репликация ДНК клеточного ядра, был назван " фаза S " (от слова synthesis). Период между фазой М и началом фазы S обозначен как фаза G1 (от слова gap - промежуток), а период между концом фазы S и последующей фазой М - как фаза G2. Период клеточного деления (фаза М) включает две стадии: митоз (деление клеточного ядра) и цитокинез (деление цитоплазмы). В свою очередь, митоз делится на пять стадий (рис.3), In vivo эти шесть стадий образуют динамическую последовательность. Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
Повторяющаяся совокупность событий, обеспечивающих деление эукариотических клеток, получила название клеточного цикла. Продолжительность клеточного цикла зависит от типа делящихся клеток. Некоторые клетки, например, нейроны человека, после достижения стадии терминальной дифференцировки прекращают свое деление вообще. Клетки легких, почек или печени во взрослом организме начинают делиться лишь в ответ на повреждение соответствующих органов. Клетки эпителия кишечника делятся на протяжении всей жизни человека. Даже у быстро пролиферирующих клеток подготовка к делению занимает около 24 ч. Клеточный цикл разделяют на стадии: Митоз - М-фаза, деление клеточного ядра. G1 -фаза период перед синтезом ДНК. S-фаза - период синтеза (репликации ДНК). G2-фаза - период между синтезом ДНК и митозом. Интерфаза - период, включающий в себя G1 -, S- и G2-фазы. Цитокинез - деление цитоплазмы. Точка рестрикции, R-point - время в клеточном цикле, когда продвижение клетки к делению становится необратимым. G0 фаза - состояние клеток, достигших монослоя или лишенных фактора роста в ранней G1 фазе.
Делению клетки (митозу или мейозу) предшествует удвоение хромосом, которое происходит в периоде S клеточного цикла (рис.1). Период обозначают первой буквой слова synthesis - синтез ДНК. С момента окончания периода S до завершения метафазы ядро содержит в четыре раза больше ДНК, чем ядро сперматозоида или яйцеклетки, а каждая хромосома состоит из двух идентичных сестринских хроматид. Во время митоза хромосомы конденсируются и в конце профазы или начале метафазы становятся различимыми при оптической микроскопии. Для цитогенетического анализа обычно используют препараты именно метафазных хромосом.
В начале анафазы центромеры гомологичных хромосом разъединяются, и хроматиды расходятся к противоположным полюсам митотического веретена. После того как к полюсам отойдут полные наборы хроматид (с этого момента их называют хромосомами), вокруг каждого из них образуется ядерная оболочка, формируя ядра двух дочерних клеток (разрушение ядерной оболочки материнской клетки произошло в конце профазы). Дочерние клетки вступают в период G1, и только при подготовке к следующему делению они переходят в период S и в них происходит репликация ДНК.
Клетки со специализированными функциями, длительное время не вступающие в митоз или вообще утратившие способность к делению, находятся в состоянии, называемом периодом G0. Большинство клеток в организме диплоидные - то есть имеют два гаплоидных набора хромосом (гаплоидный набор - это число хромосом в гаметах, у человека он составляет 23 хромосомы, а диплоидный набор хромосом - 46). В гонадах предшественники половых клеток сначала претерпевают ряд митотических делений, а затем вступают в мейоз - процесс образования гамет, состоящий из двух последовательных делений. В мейозе гомологичные хромосомы спариваются (отцовская 1-я хромосома с материнской 1-й хромосомой и т. д.), после чего в ходе так называемого кроссинговера происходит рекомбинация, то есть обмен участками между отцовской и материнской хромосомами. В результате качественно изменяется генетический состав каждой из хромосом.
В первом делении мейоза расходятся гомологичные хромосомы (а не сестринские хроматиды, как в митозе), вследствие чего образуются клетки с гаплоидным набором хромосом, каждая из которых содержит по 22 удвоенные аутосомы и одной удвоенной половой хромосоме. Между первым и вторым делениями мейоза нет периода S (рис.2, справа), а в дочерние клетки во втором делении расходятся сестринские хроматиды. В итоге образуются клетки с гаплоидным набором хромосом, в которых вдвое меньше ДНК, чем в диплоидных соматических клетках в периоде G1, и в 4 раза меньше - чем в соматических клетках по окончании периода S.
При оплодотворении число хромосом и содержание ДНК у зиготы становится таким же, как в соматической клетке в периоде G1. Период S в зиготе открывает путь к регулярному делению, характерному для соматических клеток.