РАЗВИТИЕ ЗАРОДЫШЕВЫХ ОБОЛОЧЕК ИЛИ ПРОВИЗОРНЫХ ОРГАНОВ У ПТИЦ
ВВЕДЕНИЕ
Необходимость настоящих методических разработок определяется рядом причин. Во-первых, в овладении предметом акцент переносится на самостоятельную работу студентов, в том числе во внеучебное время. Это требует вооружения студентов дополнительной литературой и методическими разработками, которые должны оптимизировать этот процесс.
Во-вторых, преподавание, эмбриологии, одной из фундаментальных дисциплин в медицинской подготовке будущего врача, искусственно расчленено и проводится на разных кафедрах, начиная с кафедр биологии, анатомии человека, гистологии и эмбриологии, акушерства и гинекологии, а также на многих других, где рассматриваются врожденные уродства и аномалии развития. Все это требует создания методических разработок, которые помогли бы студенту при изучении с помощью учебника и дополнительной литературы освоить этот довольно сложный раздел теоретической медицины. .
Решению поставленных задач должны помочь методические разработки «Основы эмбриологии». Они состоят из 3-х разделов. Первый включает дополнения и комментарии к теоретическому материалу курса. Второй — задания к проведению практических занятий и ориентировочной карты к самостоятельным действиям студентов. Третий — контрольные вопросы, контрольно-обучающие и ситуационные задачи по разделу «Эмбриология».
При подготовке настоящих мётодразработок были использованы материалы методических разработок, созданные коллективом кафедры гистологии 1-го Московского медицинского института им. И. М. Сеченова под редакцией профессора Ю. А. Афанасьева.
Раздел задач по «Эмбриологии человека» подготовлен сотрудниками кафедры гистологии педиатрического факультета 2-го Московского медицинского института им. Н. И. Пирогов а (зав.-проф. О. В. Волкова). Этим коллективам приносим нашу благодарность.
ОПРЕДЕЛЕНИЕ, ПРЕДМЕТ И ЗАДАЧИ
«Эмбриология — «наука о развитии зародыша. Под зародышевым или эмбриональным развитием понимается тот ранний период индивидуального развития организма, который проходит от
момента оплодотворения до рождения на свет (у живородящих организмов) или до вылупливания из яйцевых оболочек (у яйцекладущих животных)». (А. Г. Кнорре, 1914—1982).
Предметом изучения эмбриологии является также прогенез — предзародышевый период (формирование половых клеток) и по- слезародышевый- (постэмбриональный) или послеродовой (постна- тальный) периоды индивидуального развития. У млекопитающих: некоторые системы органов (например, половая, эндокринная) приобретают дефинитивные, т. е. окончательные, свойственные взрослому состоянию, структуры и функции спустя тот или иной промежуток времени после рождения.
Эмбриология исследует как общие закономерности индивидуального развития, проявляющиеся в развитии всех'многоклеточных животных организмов, так и частные особенности процессов индивидуального развития, характерные для представителей отдельных типов, классов или видов животных.
В связи с этим различают общую эмбриологию, рассматривающую наиболее общие вопросы и наиболее широкие закономерности индивидуального развития, и частную эмбриологию отдельных групп животных.
Значение эмбриологии заключается прежде всего в формировании общих биологических представлений на- материалистической основе. Велико значение эмбриологии как теоретической основы педиатрии, акушерства и гинекологии, тератологии.
МЕТОДЫ ИССЛЕДОВАНИЯ
Для изучения эмбрионального развития животных пользуются различными методами.
Простейший и наиболее древний из них — наблюдение и описание развития зародыша у тех или иных животных или человека. На основе этого метода возникла описательная эмбриология.
Значительно позже в эмбриологических исследованиях стал применяться сравнительный метод, создавший сравнительную эмбриологию. Ее возникновение было обусловлено накоплением фактов о развитии различных видов животных. Применение сравнительного метода открыло возможности для понимания общих закономерностей развития. С помощью сравнительного метода был открыт закон зародышевого сходства К. М. Бэром (1792—1876).
Описательная и сравнительная эмбриология стали фундаментом для развития эволюционной эмбриологии, создателями которой явились А. О. Ковалевский (1840—1901), И. И. Мечников (1845— 1916). Развитие эволюционной эмбриологии привело к открытию биогенетического закона Э. Геккелем (1834—1919), Ф. Мюллером (1821—1897), А. Н, Северцевым (1861—1936).
С развитием и усложнением экспериментальной техники воз
никло новое направление исследований — экспериментальная эмбриология. Основателями ее явились В. Ру (1850—1924), Г. Дриш (1867—1941) и Г. Шпеман (1869—1941). В основе экспериментальной эмбриологии лежит изучение последствий удаления или трансплантация различных частей зародыша, а также воздействия на зародыш или его части разных экзогенных факторов.
Современная эмбриология пользуется всеми видами световой, электронной и сканирующей микроскопии, методами прижизненного наблюдения эмбриогенеза при помощи меток, наносимых на зародыш с помощью изотопов или красителей, методами гистохимии, биохимии, радиологии, иммунологии, а также статистики
ТЕОРИИ ЭМБРИОЛОГИИ
. Как и любая другая наука, эмбриология на каждом этапе развития была вооружена собственной теорией.
С именем Гиппократа (460—377 год до н. э.) и Аристотеля (384—322 год до н. э.) связывают не только определенные знания в области развития организмов, но и создание первых научных эмбриологических представлений.
Согласно теории Гиппократа, плод образуется через смешение мужского и женского семени. Он считал, что все члены организма 1возникают независимо друг от друга. Ни один из них не возникает раньше или позже другого.
Эта идея вновь возродилась и стала господствовать в течение XVII—XVIII вв. Она получила название теории преформизма. Согласно этой теории каждый зародыш с самого начала является уже вполне сформированным, имеющим все части организма, которому остается только расти. Наиболее крайние и наивные преформисты представляли себе, что каждый организм, в том числе и человеческий, заключает в себе громадное количество поколений зародышей, вложенных друг в друга в теле прародителей со времени сотворения мира.
Между представителями двух направлений преформистов велись горячие споры. Анималькулисты или сперматики считали, что зародыш преформирован в сперматозоиде. Овисты думали, что зародыши в миниатюрном виде находятся не в мужских половых клетках, а в яйце.
Сторонниками префоризма были многие выдающиеся ученые XVII и XVIII века: А. Левенгук (1632—1723), Я. Сваммердам (1637—1680), М. Мальпиги (1628—1694), А. Галлер (1708—1777), Ш. Бонне (1720—1793).
Аристотель впервые сформировал теорию, эпигенеза. Согласно Аристотелю, части зародыша не заложены в невидимо малых размерах в «семенах вещей» и не возникают все одновременно, а растут и развиваются в определенной последовательности. Это положение дополнялось виталистическим взглядом о том, что материал материнского зачатка приобретает форму будущего организма под влиянием нематериальных сил — «энтелехий», заложенных в семени отца.
Виталистический взгляд Аристотеля в XVII веке был возрожден У. Гарвеем (1578—1657), предложившим и термин «эпигенез». У. Гарвей выдвинул принцип «все из яйца». Причем у животных, у которых ему не удалось обнаружить яиц, он допускал их зарождение из «разлагающихся веществ».
Однако, в обеих этих интерпретациях теория эпигенеза успеха не имела, и во второй половине XVII и в первой половине XVIII века господствовала теория преформизма.
Теория преформизма вновь возродилась во второй половине XVIII века в связи с распространением учения о всемирном тяготении и влиянии его на мир химических и биологических процессов.
Обоснование эпигенезу дал К. Ф. Вольф (1733—1794). Он утверждал, что лишь в результате поступления извне питательных соков в зачатке образуются клетки и сосуды, из которых затем формируются органы будущего зародыша. К. Ф. Вольф ссылался на наличие «существенной силы», которая обуславливает наступление и продвижение питательных веществ, необходимых для образования частей зародыша.
Борьба теорий преформизма и эпигенеза стала особенно ожесточенной в конце XIX века в связи с развитием экспериментальной эмбриологии и, так называемой механики развития, претендовавших на каузальное объяснение онтогенеза.
Однако, в те времена речь шла уже не о наличии в зачатковых клетках готового миниатюрного зародыша, а о химических и морфологических структурах, предопределяющих развитие организма. Эти положения дополнялись виталистическим взглядом о том, что материал материнского зачатка приобретает форму будущего организма под влиянием нематериальных сил — «энтелехий», заложенных в семени отца.
Полемика между сторонниками эпигенеза и преформизма велась также вокруг проблемы соотношения внутренних и внешних факторов наследственности в эволюции. Представители эпигенеза пытались обосновать свои взгляды с ссылкой на явления наследования приобретенных признаков, на возможность влияния внешней среды на онто- и филогенез, т. е. опирались на различные ламарксистские теории. Однако, многочисленные попытки подтвердить эти представления экспериментально оказались безуспешными. Вместе с тем успехи молекулярной биологии и генетики во второй половине XX столетия доказали наличие в зачатковых клетках предетермированных химических и биологических структур (молекул ДНК), которые определяют воспроизведение основных видов и индивидуальных особенностей развивающегося организма.
СОВРЕМЕННАЯ ЭМБРИОЛОГИЯ
Ставит своей задачей выявление движущих сил эмбрионального формообразования, вскрытие генетических и цитологических основ, клеточной, тканевой и органной дифференцировки.
В 20—40 годы XX века большую роль в развитии эмбриологии сыграли работы Г. Шпемана и его школы по изучению влияния одних частей зародыша на другие; были выведены понятия «индуктор», «организатор».
Д. П. Филатов (1876—1943) и другие советские исследователи развили учение Г. Шпемана и внесли в него существенные поправки.
Д. П. Филатов связал экспериментальную эмбриологию с эволюционным учением и сформулировал понятие о формообразовательном аппарате, т. е. тех частей зародыша, взаимодействие которых приводит к осуществлению определенных этапов развития.
Советские ученые сделали существенный вклад и в понимание закономерностей индивидуального развития. Н. И. Кольцов впервые произвел попытку синтеза эмбриологии и генетики. П. Г. Светлов предложил оригинальный вариант теории «критических периодов» в развитии организма. Б. П. Токин начал исследование соматического эмбриогенеза, т. е. изучение развития организмов из соматических клеток.
В настоящее время процесс развития понимается как реализация тех возможностей, которые заложены в «наследственном коде» половых клеток. Хотя для осуществления этих процессов развития необходимо наличие определенных условий среды, они неизменно реализуются путем самодифференцировки на основе тех видоспецифических закономерностей, которые были исторически «выработаны и закреплены на протяжении эволюции органического мира.
ПОЛОВЫЕ КЛЕТКИ ИЛИ ГАМЕТЫ
Сперматозоиды позвоночных имеют жгутиковую форму и обладают способностью к активному движению. В них различают головку, шейку, тело и хвост.
Головка сперматозоида содержит плотное ядро, окруженное тонким слоем цитоплазмы. В ядре содержится гаплоидный набор хромосом, содержащих ДНК, РНК» протамины, гистоны. Сверху головка сперматозоида покрыта чехликом, в котором располагается плотная гранула — акросома. Чехлик и акросома являются производными пластинчатого комплекса Гольджи. Акросома содержит фермент гиалуронидазу, способную растворять оболочки, покрывающие яйцеклетки.
Шейка сперматозоида содержит проксимальную центриоль, играющую важную роль в делении оплодотворенного яйца, и дистальную, от которой отходит осевая нить, проходящая через тело и хвост. Тело сперматозоида содержит митохондрии, которые по спирали окружают осевую нить, а также вторую половину дистальной центриоли. В теле находятся запасы гликогена.
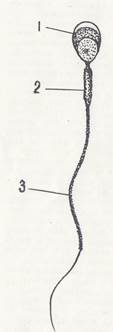
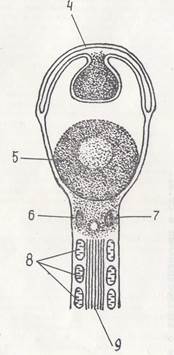
Рис. 1. Строение сперматозоида .
1—головка; 2 — шейка; 3 — хвост; 4 — акросома, 5 — ядро; .6 дистальная центриоля; 7 — проксимальная центриоля; 8 — митохондрии; 9 дипломакро-фибриллы центральные и периферические
Хвост сперматозоида содержит осевую нить, которая окружена небольшим количеством цитоплазмы, содержащей фермент адено зинтрифосфатазу. Скорость движения сперматозоида 2—3 мм в минуту (рис. 1) .
Женская половая клетка — яйцеклетка — имеет округлую форму и не обладает способностью к самостоятельному движению. Яйцеклетка содержит крупное ядро с гаплоидным набором хромосом, ядрышко и ооплазму, в которой располагается питательный материал в виде желтка. Величина яйцеклетки зависит от количества отложенного в ооплазме желтка.
Размеры яйцеклеток млекопитающих имеют диаметр 50—150 микрометров. Яйцеклетка содержит слаборазвитую цитоплазматическую сеть, митохондрии, пластинчатый комплекс Гольджи. В зрелой яйцеклетке отсутствует центросома. По периферии располагаются кортикальные гранулы, содержащие кислые мукополисахариды. Яйцеклетка покрыта первичной оболочкой, которая образуется путем затвердения наружного слоя цитоплазмы в ходе оогенеза, и вторичной оболочки, образованной фолликулярными клетками и называемой блестящей оболочкой. Ее наружная часть, состоящая из кислых мукополисахаридов, секретируется фолликулярными клетками. Внутренняя зона, состоящая из нейтральных мукополисахаридов, образуется ооцитом. Фолликулярные клетки выполняют трофическую функцию, доставляя в яйцеклетку питательные вещества.
ТИПЫ ЯЙЦЕКЛЕТОК
Классификация яйцеклеток основана на количестве и распределении желтка, содержащегося в ооплазме. Желток — совокупность включений, состоящих из питательных веществ. Яйцеклетки с небольшим количеством желтка, распределенном равномерно по всей цитоплазме, называются изолецитальными. Они характерны для ланцетника и морских ежей (рис. 2а).
Яйцеклетки со средним количеством желтка; распределенном на нижнем вегетативном полюсе яйца, называются умеренно телолецитальными. Цитоплазма концентрируется на верхнем анимальном полюсе. Такие яйцеклетки характерны для амфибий (рис. 26).
Яйцеклетка, содержащая большое количество желтка, заполняющего почти всю яйцеклетку, называется резко телолецитальной. Примером являются яйцеклетки акуловых, костистых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих (ехидна, утконос).
У млекопитающих и человека в связи с внутриутробным развитием и питанием зародыша за счет материнского организма, яйцеклетка в процессе эволюции стала изолецитальной, возникнув из телолецитальной. Такие яйцеклетки, называются вторично х изолецитальными.



ЭМБРИОГЕНЕЗ
Эмбриогенез слагается из следующих сменяющих друг друга процессов и стадий:
Процессы и стадии эмбриогенеза

ОПЛОДОТВОРЕНИЕ
Оплодотворением называется процесс слияния мужской и женской гамет в одну клетку, в результате которого образуется новый организм — зигота.
В процессе оплодотворения различаем два этапа: проникновение сперматозоида через оболочки яйцеклетки и слияние ядер.
Сперматозоиды, приближаясь к яйцу, выделяют фермент гиалуронидазу, под действием которого мукополисахариды, скрепляющие окружающие яйцо фолликулярные клетки, разрушаются. Яйцо освобождается от оболочки, образованной фолликулярными клетками. Один из сперматозоидов проникает через оболочку яйцеклетки. В цитоплазму проникает головка, шейка и тело. Кортикальные гранулы набухают и превращаются в вакуоли, которые лопаются, а их водянистое содержимое образует под желточной оболочкой перивителлиновое пространство. Быстрое образование перивителлинового пространства и утолщение желточной оболочки приводит к появлению оболочки оплодотворения, которая препятствует внедрению других сперматозоидов. Сперматозоиды вносят в яйцеклетку центросому, благодаря которой происходит дальнейшее деление оплодотворенного яйца.
Второй этап процесса оплодотворения — слияние мужского и женского пронуклеусов (ядер, содержащих гаплоидный набор хромосом) и образование единого ядра—синкариона (рис. 3).
Таким образом, основными моментами оплодотворения являются:
1. Внедрение спермия в яйцеклетку.
2. Образование оболочки оплодотворения.
3. Слияние пронуклеусов и образование синкариона.
Биологическое значение оплодотворения состоит в том, что в зиготе восстанавливается нормальный для данного вида диплоидный набор хромосом (для человека 46), происходит передача наследственных свойств новому организму, половые хромосомы определяют пол организма; активизируется синтез АТФ, уъинуь ваются обменные процессы, протеазы расщепляют запасы желтка, возникает синтез новых белков, начинает осуществляться программа развития новой особи.


Рис. 3. Процесс оплодотворения 1 яйцеклетка; 2 — сперматозоид; 3 — ядро яйцеклетки; 4 — ядро сперматозоида; 5 — центросома; 6 — синкарион
ДРОБЛЕНИЕ
Дроблением называется процесс, в результате которого из зиготы путем последовательных делений образуется многоклеточный организм. Дробление происходит путем митоза. Образовавшиеся новые клетки — бластомеры не растут, а с каждым новым делением становятся все более мелкими. Увеличивается число бластомеров, но в сумме их масса не превышает массы зиготы. Дробление характерно для всех хордовых животных, но в зависимости от количества и распределения желтка у различных групп животных оно происходит по-разному. Желток является питательным материалом будущего зародыша, но он замедляет деление клеток. Чем больше желтка в цитоплазме зиготы, тем медленнее делится эта часть зародыша.
Изолецитальные яйцеклетки содержат мало желтка, который распределен примерно равномерно. Эти клетки делятся целиком на одинаковые бластомеры и количество их после каждого этапа деления увеличивается вдвое. Такое дробление называется полным (голобластическим) и равномерным. При этом первая и вторая борозды дробления идут меридиально, перпендикулярно одна к другой. Третья—экваториально. Затем борозды дробления чередуются, в результате возникает зародыш из 128 бластомеров (пример, ланцетник).
Умеренно телолецитальные яйцеклетки (пример, амфибии) делятся. полностью. Первые две борозды дробления происходят меридиально, третья — выше экватора, т. е. тангенциально. На анимальном полюсе, где желтка меньше, дробление идет быстрее и получаются мелкие бластомеры (микромеры), на вегетативном полюсе, где желтка много, дробление идет медленнее, возникают крупные бластомеры (макромеры). Такое дробление будет полным, но не равномерным.
Резкотелолецитальные яйцеклетки (пример, птицы), дробятся частично (меробластически). Делится цитоплазма анимального полюса бластулы, расположенная в виде диска. Вегетативный полюс, заполненный желтком, не делится. Такое дробление называется дискоидальным.
Вторичнотелолецитальные яйцеклетки (пример, млекопитающие в том числе и человек) дробятся полностью, неравномерно и образуют не одинаковые по размерам и свойствам (неравноценные) (
бластомеры. Крупные бластомеры (темные) дадут в будущем эмбриобласт, т. е. материал для развития зародыша, мелкие (светлые) — дадут трофобласт, материал для одной из зародышевых оболочек. Дробление у млекопитающих и человека получило- название полного, неравного и неравноценного.
Биологическое значение дробления состоит в том, что с помощью этого процесса происходит переход к многоклеточной форме организации зародыша. В ходе дробления увеличивается общее- содержание и синтез ДНК и некоторых РНК, общая суммарная поверхность клеток зародыша, ядерно-цитоплазматическое отношение, происходит разделение некоторых веществ цитоплазмы, происходит подготовка к региональной дифференцировке.
БЛАСТУЛА
В результате дробления зародыш становится многоклеточным внутри себя имеет полость и переходит на стадию бластулы. Бластула имеет стенку—бластодерму, полость — бластоцель, заполненную жидкостью.
При полном и равномерном дроблении (ланцетник, морской, еж) образуется целобластула. В ней следует различать дно, представленное бластомерами вегетативного полюса, крышу, образованную бластомерами анимального полюса, и краевую зону, расположенную между полюсами.
При полном неравномерном дроблении (амфибии) образуется: амфибластула, где бластодерма многослойная, а бластоцель расположена эксцентрично, ближе к анимальному полюсу. Крыша амфибластулы тонкая, так как состоит из микробластомеров, дно бластулы образовано наполненными желтком бластомерами.
При частичном дискойдальном дроблении (акуловые, костистые рыбы, пресмыкающиеся, птицы, низшие млекопитающие) образуется дискобластула. Она представляет собой зародышевый диск, расположенный на желтке. Зародышевый, диск соответствует крыше и краевой зоне бластулы, желточная масса дну, а узкая щель между ними представляет бластоцель.
При полном неравномерном и неравноценном дроблении (млекопитающие, человек) образуется плотный клеточный шар, называемый стерробластулой. В ней возникает полость и зародыш превращается в бластоцисту, в которой различают стенку — трофобласт и скопление темных бластомеров на внутренней поверхности: трофобласта—эмбриобласт (рис. 4).

Рис. 4. Дробление изоледитальной яйцеклетки ланцетника
1 — бластомеры
ГАСТРУЛЯЦИЯ
Под гаструляцией понимают процесс превращения бластулы (двухслойного зародыша, состоящего из двух зародышевых листков, наружного — эктодерма и внутреннего — энтодерма) в гаструлу. Этот процесс приводит к обособлению третьего зародышевого листка — мезодермы.
Гаструляция у животных всех классов представляет собой существенную структурную перестройку зародыша, осуществляемую путем клеточных движений и перемещений с образованием избирательных связей. Гаструляция обеспечивает установление основного структурного плана в виде 3-зародышевых листков и дальнейшее развитие зародыша. Поскольку гаструляция является продолжением изменений, начавшихся на стадии бластулы, то различным типам бластул соответствуют различные типы гаструл. Поэтому гаструляция у разных животных протекает различно.
Но как бы ни были разнообразны переходы бластулы в гаструлу, сущность процессов заключается в том, что происходит перемещение зародышевого материала и обособление 3-зародышевых листков.
Перемещение клеточного материала при гаструляции осуществляется путем иммиграции, инвагинации, эпиболии, деляминации.
ВИДЫ ГАСТРУЛЯЦИИ
По И. И. Мечникову наиболее древним способом гаструляции является иммиграция. Этот способ заключается в активном выселении части бластомеров в бластоцель. Такая иммиграция называется униполярной. Выселение бластомеров может происходить по всей поверхности бластодермы. В таком случае иммиграция называется мультиполярной. Она представляет редкое явление. Выселившиеся в бластоцель бластомеры образуют первичный внутренний зародышевый листок (энтодерму), а оставшиеся на месте бластомеры образуют первичный наружный зародышевый листок (эктодерма). Этот способ распространен у низших животных— рептилий, птиц, млекопитающих. Сама форма зародыша при иммиграции называется иммиграционной гаструлой.
Инвагинация — впячивание, наиболее простой способ гаструляции. Заключается во впячивании бластодермы вегетативного полюса вглубь бластоцеля. Появившееся впячивание все больше углубляется и в конце концов доходит до анимального полюса. В результате зародыш из однослойного пузырька превращается в двуслойный. Его стенка состоит из первичного наружного и первичного внутреннего листков. Вследствии впячивания вегетативного полюса бластоцель вытесянется. Образуется новая полость — полость первичной кишки или гастроцель. Она сообщается с внешней средой отверстием — первичным ртом или бластопором. Края бластопора называют губами. Различают дорзальную и вентральную губы бластопора. Такой способ гаструляции имеет место" у ланцетника, иглокожих, низших хордовых.
Эпиболия — обрастание, способ гаструляции, при котором быстро делящиеся мелкие бластомеры анимального полюса (микромеры) наползают на крупные бластомеры вегетативного полюса
(макромеры), не способные к перемещению вследствие загруженности желтком. Первые образуют эктодерму, а вторые — энтодерму. Такая гаструляция характерна для амфибий.
Деляминация —расщепление, способ гаструляции, заключающийся в параллельном расслоении поверхностного слоя бластомеров на два: первичный наружный, первичный внутренний. Такой способ гаструляции наблюдается у некоторых членистоногих и у высших позвоночных.
У некоторых животных гаструляция может осуществляться комбинацией двух или более различных способов.
ГАСТРУЛЯЦИЯ У ЛАНЦЕТНИКА
Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный— из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша — образование третьего зародышевого листка—мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку— зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж — хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок — мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму — внутренний зародышевый листок (рис. 5).

ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью — бластоцель. Крыша этой бластулы представлена бластодиском, дно — нераздробившейся массой желтка. Бластоцель — подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость — гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение— первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост — зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы — третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой

Рис.%6. Развитие комплекса осевых органов у птиц 1—эктодерма; 2 —нервный желобок; 3 — нервная трубка; 4 — амнион; 5 — сомиты; 6 — париетальный листок сплахнотома; 7 — висцеральный листок спланхнотома; 8 — хорда; 9 - дерматом; 10 — миотом; 11 — склеротом; 12 — нефротом; 13 — аорта; 14 — энтодерма; 15 — амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
НЕЙРУЛЯЦИЯ
Процесс, обеспечивающий переход зародыша от стадии гаструлы на стадию нейтрулы, получил название нейруляции. При этом, помимо нервной трубки, идет образование хорды и сомитов. Эта триада называется органами спинного комплекса.
В период нейруляции по средней линии на спинной стороне зародыша в результате размножения клеток образуется утолщение — нервная пластинка. Из этой же нейральной эктодермы формируются гребневидные складки — нервные валики, по бокам ограничивающие нервную пластинку. Нервные валики появляются сначала на переднем конце тела зародыша, а затем в средних и задних частях его.
Клетки нервной пластинки, размножаясь погружаются вовнутрь зародыша, образуя нервный желобок. Затем края этого желобка смыкаются и* возникает нервная трубка, содержащая полость — невроцель. Широкая передняя часть нервной трубки в дальнейшем даст структуры головного мозга, а более узкая туловищная часть — спинной мозг. Нервные валики приподнимаются над дорзальной поверхностью нервной трубки, сближаются друг с другом, формируют непарную ганглиозную пластинку. Эктодерма погружает под себя ганглиозную пластинку и нервную трубку, и в дальнейшем превращается в поверхностный хлой кожи--эпидермис. После смыкания кожной эктодермы ганглиозная пластинка располагается между ней и нервной трубкой и представляет собой тяж клеток, называемый нервным гребнем. Это временное скопление клеток, которое быстро распадается после возникновения.
Большинство клеток нервного гребня мигрирует по направлению к вентральной стороне зародыша.
Из клеток нервного гребня в дальнейшем образуются меланоциты — клетки, содержащие пигмент меланин, придающий коже окраску, а также, вероятно, клетки диффузной нейроэндокринной системы.
В самом начале нейруляции до появления нервной трубки по средней линии в передней половине зародышевого щитка из общего зачатка хордо-мезодермы обособляется хорда, по бокам которой располагается парахордальная мезодерма. Ее подразделяют на дорзальную и вентральную.
Дорзальная мезодерма в передне-заднем направлении -сегментируется на сомиты, в которых в дальнейшем выделяют три участка, получившие соответственно названия: дерматом, миотом и склеротом.
Вентральная мезодерма разделяется параллельно поверхности тела на два листка: париетальный, прилежащий к эктодерме, и висцеральный, прилежащий к энтодерме. Между этими листками появляется узкая щель — вторичная полость тела или целом.
Между дорзальной и вентральной мезодермой, объединяя их,

Рис. 7. Провизорные органы у птиц 1 — зародыш; 2 — амнион; 3 — серозная оболочка; 4 — аллантоис; 5 — желточный мешок; 6 — воздушная камера; 7 — белок; 8 — скорлупа
обособляются сегментные ножки или нефротомы, материал из которого в дальнейшем даст некоторые структуры мочеполовой системы. Общий план строения зародыша на поперечном срезе представлен на приводимой схеме (рис. 7).
ГИСТОГЕНЕЗ
Эмбриональный гистогенез — процесс возникновения тканей из малодифференцированных клеток эмбриональных зачатков. Эмбриональные зачатки (кожная эктодерма, кишечная энтодерма, нервная трубка, ганглиозная пластинка, миотом и др.) к концу нейруляции представлены сравнительно небольшими группами малодифференцированных клеток и не имеют тканевого строения. В отличие от тканей, в составе которых присутствуют специализированные клетки и могут быть структуры, не имеющие клеточного строения (симпласты, межклеточное вещество и др.), эмбриональные зачатки имеют вначале только клеточный состав.
Эмбриональная гистогенез — сложный процесс, который можно с известной степенью условности расчленить на 3 группы более простых состовляющих процессов. К первой группе можно отнести процессы изменения числа клеток эмбрионального зачатка за счет их размножения (а также гибели клеток). Ко второй группе — процессы изменения пространственного расположения клеток за счет их перемещения. К третьей группе — процессы дцфференциации клеток за счет изменения их структуры, размеров и_функции. В течение дифференциации клетки эмбрионального зачатка постепенно приобретают структурные и функциональные свойства, характерные для той или иной ткани. Параллельно с дифференциацией осуществляется повышение степени интегрировянности развивающихся тканей за счет взаимодействия клеток, составляющих эти ткани.
Перечисленные процессы необходимы для образования 4 групп тканей: эпителиальных, соединительных, мышечных, нервной. Частные закономерности формирования каждой из тканей будут рассмотрены в курсе общей гистологии. Здесь перечислим эмбриональные зачатки и развивающиеся из них ткани.

|

РАЗВИТИЕ ЗАРОДЫШЕВЫХ ОБОЛОЧЕК ИЛИ ПРОВИЗОРНЫХ ОРГАНОВ У ПТИЦ
Зародышевые оболочки или провизорные органы — временные структуры. Они образуются и функционируют лишь в эмбриональном периоде, создавая благоприятные условия для развития зародыша. В конце эмбрионального периода они теряют связь с зародышем и погибают. К провизорным органам относят;
1. Желточный мешок.
2. Амнион.
3. Серозная оболочка
4. Аллантоис.
Образование и функции желточного мешка
У птиц желточный мешок начинает формироваться на стадии ранней гаструлы. В этот период развития зародыш имеет вид диска, распластанного на желтке. Зародышевый диск состоит из двух клеточных тяжей: наружного — эктодермального и внутреннего— энтодермального. Клетки энтодермы, прилегающие к желтку, выделяют протеолитические ферменты, которые расщепляют последний и всасывают продукты его расщепления. Тем самым обеспечивается питание зародыша птиц на ранних этапах его развития. В результате под зародышем образуется полость, свободная от желтка.
При попадании зародыша в условия инкубации потребность его в притоке питательных веществ резко увеличивается. Образованием туловищных складок, формирующихся от головного к хвостовому концу зародыша, отделяется тело от остатков желтка. Туловищную складку образуют все зародышевые листки: энтодерма, висцеральная и париетальная вентральная мезодерма, эктодерма. Свободная от желтка полость, прилежащая к телу зародыша и отграниченная энтодермой, образует кишечную трубку. Нижерасположенный желток, отграниченный энтодермой и висцеральным листком вентральной мезодермы, сообщается с полостью кишечной трубки желточным стебельком. Эта структура, состоящая из энтодермы и висцерального листка вентральной мезодермы и охватывающая желток, получила название желточного мешка. В стенке последнего в дальнейшем образуются кровеносные сосуды, куда, будут поступать продукты гидролиза желтка, всасываемые энтодермой желточного мешка. Помимо сказанного, в стенке желточного мешка возникнут первые клетки крови, сюда будут мигрировать первичные половые клетки с последующим их перемещением в половые железы (рис. 7).
Образование, строение и функции амниотических складок, амниона и серозной оболочки
Одновременно с туловищной складкой вокруг тела зародыша закладываются амниотические складки. Они располагаются вентрально и параллельно туловищным складкам, но затем верхушки их направляются кверху и к центру, охватывая тело зародыша с боков и сзади. Эти складки образованы снаружи внезародышевой эктодермой, внутри — париентальным листком вентральной мезодермы.
Амниотические складки закладываются на головном конце зародыша, образуя как бы капюшон над ним, затем распространяются к заднему концу и срастаются над зародышем. При этом эктодермальные листки срастаются с эктодермальными, мезодер-

Рис. 8. Развитие зародыша человека 1 трофобласт; 2 —• эмбриобласт; 3 —эктодерма; 4 —энтодерма; 5 —зародышевый щиток; 6 — амниотический пузырек; 7 — желточный пузырек; 8 —
внезародышевая мезодерма
мальные — с мезодермальными. В результате этого вокруг тела зародыша образуются две оболочки. Внутренняя — амниотическая, наружная — серозная.
Амниотическая оболочка изнутри представлена внезародышевой эктодермой, снаружи — париетальным листком вентральной мезодермы. Амниотическая оболочка вначале вплотную прилегает к телу зародыша, затем между ними возникает амниотическая полость, которая быстро заполняется белковой жидкостью.
Тем самым, функция амниотическях складок реализуется образованием амниона, в водной среде которого как и у низших позвоночных идет развитие зародыша, защищенного от высыхания и других повреждающих факторов внешней среды.
Серозная оболочка, расположенная над амниотической, также состоит из двух листков. Эктодермальный — обращен наружу, висцеральный листок мезодермы располагается под эктодермальным. Серозная оболочка совместно с аллантоисом участвует в газообмене и резорбции желтка.
Если дорзальные концы амниотических складок образуют амниотическую и серозную оболочки, то вентральные отделы этих складок растут кпёреди и с боков окутывают желточный мешок и срастаются под ним (рис. 7).

Рис. 9. Имплантация зародыша в слизистую оболочку матки 1—эмбриобласт; 2— трофобласт; 3 — ворсинки трофобласта; 4 — эпителий эндометрия; 5 г— собственная пластинка эндометрия
Образование и функции аллантоиса
Аллантоис образуется в виде колбасовидного выроста боковой стенки первичной задней кишки. Он состоит из кишечной энтодермы и висцерального листка вентриальной мезодермы (рис. 7).

Рис. 10. Строение плаценты человека 1 — амниотичеСкий эпителий; 2— пупочный канатик; 3 — пупочная вена; 4— пупочные артерии; 5 — эпителий ворсин плаценты; 6 — капиллярные петли ворсин плаценты; 7 — кровеносные лакуны; 8 — децидуальная оболочка матки; 9 —
миометрий
В результате роста аллантоис постепенно внедряется между желточным мешком, амнионом и серозной оболочкой, располагаясь вокруг зародыша. Своим мезодермальным листком аллантоис прилежит к мезодерме серозной оболочки и срастается с ней. Из стенки задней кишки по мезодермальному листку в аллантоис врастают кровеносные сосуды, которые проникают по всему аллан- тоису, создавая сосудистую связь последнего с зародышем.
На границе с воздушной камерой яйца сосуды аллантоиса образуют, густую сеть капилляров, прилегающих к серозной оболочке.В этом месте происходит газообмен между кровью зародыша и воздушной камерой. Это основная функция аллантоиса — функция газообмена.
Вторая функция аллантоиса — выделительная: из сосудов аллантоиса через их стенку в просвет последнего выделяются конечные продукты белкового обмена, в частности, мочекислые соли.
Третья функция аллантоиса — трофическая: аллантоис и серозная оболочка образуют выросты, наподобие ворсинок, которые внедряются в белок, растворяют его и продукты гидролиза белка всасываются в сосуды аллантоиса и переносятся кровью к телу зародыша.
После использования всего белка, что совпадает с концом эмбрионального развития, аллантоис и прилегающая к нему серозная оболочка редуцируются, высыхают. Желточный стебелек также высыхает. Цыпленок начинает дышать воздухом, который находится внутри воздушной камеры и сразу выклевывается из яйца.