ПРОИСХОЖДЕНИЕ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
Блок 1. Строение клетки . Вопросы 2,3, 4.
Клеточная теория Т. Шванна и М. Шлейдена, ее основные положения. Современное состояние клеточной теории.
Типы клеточной организации. Строение про- и эукариотических клеток. Гипотезы происхождения эукариотических клеток (симбиотическая, инвагинационная).
Клеточная оболочка, ее структуры. Молекулярная организация и функции биологической мембраны. Виды транспорта веществ.
Клеточную теорию сформулировали впервые в 1838 году ботаник Матиас Шлейден и зоолог Теодор Шванн. Но, как и все великие открытия, теория появилась не внезапно. Свою роль здесь сыграли целые поколения ученых, которые по кирпичикам собирали её основу. Среди них следует отметить Антонио ванн Левенгука, который впервые описал простейшие организмы, сперматозоиды человека и клетки крови; Марчелло Мальпигии, увидевшего капилляры, соединяющие артерии и вены; Броуна, который в 1833 году описал клеточное ядро. Но переломным моментом стало положение Рудольфа Вирхова, который смог доказать, что клетки возникают не из первичного вещества, как считали Шлейден и Шванн, а из клеток. Это открытие повлекло за собой исследования Флеминга, описавшего в 1880 году деление клеток и последовательность процессов происходящих в митозе.
ОСНОВНЫЕ ПОЛОЖЕНИЯ СОВРЕМЕННОЙ КЛЕТОЧНОЙ ТЕОРИИ.
Клетка – элементарная структурная, функциональная и генетическая единица живого. 2) Клетки растений, животных и грибов сходны по строению, химическому составу, обмену веществ. 3) Клетка – функциональная единица живого. Сходные по строению и выполняемым функциям клетки объединяются в ткани, ткани – в органы и системы органов, которые формируют целостный организм. Это обеспечивает дискретность и целостность организма. 4) Клетка – единица развития всего живого. Любой организм начинает свое индивидуальное развитие из одной клетки – зиготы. 5) Новые клетки образуются в результате деления материнской клетки. 6) Клетка имеет мембранный принцип строения. 7) Ядро является главным компонентом эукариотической клетки, хранителем генетической информации и регуляторным механизмом. 8) Сходство строения клеток растений и животных доказывает общность их происхождения. 9) Дифференциация клеток лежит в основе усложнения строения живых организмов.
Положения клеточной теории Шванна и Шлейдена с дополнением Вирхова.
· Клетка — основная единица строения и развития всех живых организмов, является наименьшей структурной единицей живого.
· Клетки всех организмов (как одно-, так и многоклеточных) сходны по химическому составу, строению, основным проявлениям обмена веществ и жизнедеятельности.
· Размножение клеток происходит путем их деления (каждая новая клетка образуется при делении материнской клетки).
Среди всего многообразия ныне существующих на Земле организмов выделяют две группы: вирусы и фаги, не имеющие клеточного строения; все остальные организмы представлены разнообразными клеточными формами жизни. Различают два типа клеточной организации: прокариотический и эукариотический.
| Характеристика | Прокариоты | Эукариоты |
| Размеры клеток | Диаметр в среднем составляет 0,5-5 мкм | Диаметр обычно до 40 мкм; объем клетки, как правило, в 1000-10000 раз больше, чем у прокариот |
| Форма | Одноклеточные или нитчатые | Одноклеточные, нитчатые или истинно многоклеточные |
| Генетический материал | Кольцевая ДНК находится в цитоплазме и ничем не защищена. Нет истинного ядра или хромосом. Нет ядрышка | Линейные молекулы ДНК связаны с белками и РНК и образуют хромосомы внутри ядра. Внутри ядра находится ядрышко |
| Синтез белка | 70S-рибосомы и мельче. Эндоплазматического ретикулума нет. (Синтез белка характеризуется и многими другими особенностями, в том числе и чувствительностью к антибиотикам; например, развитие прокариот ингибируется стрептомицином.) | 80S-рибосомы (крупнее). Рибосомы могут быть прикреплены к эндоплазматическому ретикулуму |
| Органеллы | Органелл мало. Ни одна из них не имеет оболочки (двойной мембраны). Внутренние мембраны встречаются редко; если они есть, то на них обычно протекают процессы дыхания или фотосинтеза | Органелл много. Некоторые органеллы окружены двойной мембраной, например ядро, митохондрии, хлоропласты. Большое число органелл ограничено одинарной мембраной, например аппарат Гольджи, лизосомы, вакуоли, микротельца эндоплазматический ретикулум и т.д. |
| Клеточные стенки | Жесткие, содержат полисахариды и аминокислоты. Основной упрочняющий компонент - муреин | У зеленых растений и грибов клеточные стенки жесткие и содержат полисахариды. Основной упрочняющий компонент клеточной стенки растений - целлюлоза, у грибов - хитин |
| Жгутики | Простые, микротрубочки. Находятся вне клетки (не окружены плазматической мембраной). Диаметр 20 нм | Сложные, с расположением микротрубочек типа 9+2. Располагаются внутри клетки (окружены плазматической мембраной). Диаметр 200 нм |
| Дыхание | У бактерий происходит в мезосомах; у сине-зеленых водорослей - в цитоплазматических мембранах | Аэробное дыхание происходит в митохондриях |
| Фотосинтез | Хлоропластов нет. Происходит в мембранах, не имеющих специфической упаковки | В хлоропластах, содержащих специальные мембраны, которые обычно уложены в ламеллы или граны |
| Фиксация азота | Некоторые обладают такой особенностью | Ни один организм не способен к фиксации азота |
ПРОИСХОЖДЕНИЕ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0—1,4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение.Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней. Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0».Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания. Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов).Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот. Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Биологические мембраны
Существует несколько моделей строения биомембраны.
1. Бутербродная. (послойное расположение молекул белков, которые залегают снаружи, а липиды внутри)
2. Модель липопротеинового коврика (мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно-гидрофобных взаимодействий.)
В настоящее время общепринята жидкостно-мозаичная модель строения мембраны. Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде. Помимо липидов в состав мембраны входят белки (в среднем 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2)полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).
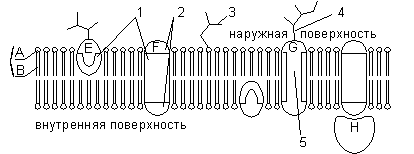
Строение мембраны: А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Функции мембран
Мембраны выполняют такие функции:
1. отделение клеточного содержимого от внешней среды,
2. регуляция обмена веществ между клеткой и средой,
3. деление клетки на компартаменты («отсеки»),
4. место локализации «ферментативных конвейеров»,
5. обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
6. распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ. Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na+, K+, Ca2+, Cl-); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na+/К+-насос (натрий-калиевый насос) (Концентрация К+ внутри клетки должна быть значительно выше, чем за ее пределами, а Na+ — наоборот), 2) эндоцитоз(процесс поглощения клеткой крупных частиц и макромолекул), 3) экзоцитоз (процесс, обратный эндоцитозу: выведение различных веществ из клетки).