ЭКСПЕРИМЕНТАЛЬНАЯ ЭМБРИОЛОГИЯ
Цель экспериментальной эмбриологии – изучение причин развития. Эксперимент ставится для того, чтобы ответить на вопрос: какие движущие силы определяют развитие? Поэтому экспериментальная эмбриология тесно связана с созданием теорий, объясняющих процесс эмбрионального развития.
Начало экспериментальных исследований относится к 1883-1869 г., когда Эрнст Геккель и Вильгельм Ру при исследовании эмбрионального развития амфибий обнаружили, что возможно развитие целых организмов из изолированных бластомеров. Несколько позже это явление было обнаружено у большинства беспозвоночных и позвоночных животных – Дриш, Шлеман, Герличка, Конклин, Шмидт, Зейдель и другие (явление однояйцевых близнецов).
80-е г. 19 столетия – годы рождения механики развития. Суть принципа методов исследований в механике развития выражалась в расшифровке причинно-следственных связей между отдельными моментами в развитии особей. То есть взаимоотношения между частями особи выражались как связи в каузальной матрице. Каузальная матрица была настолько конкретной, что могла бы быть наложена на ovo, зародыш или имаго. Считалось, что причины развития заложены в определенные части яйца или зародыша. Но применение этого метода, как известно, принесло определенный успех. Так, было открыто явление первичной эмбриональной индукции Г. Шлеманом. Но вскоре зашли в тупик, т.к. появились факты, по этой схеме не объяснимые: факт наличия эмбриональных регуляций.
Ганс Дриш – развитие не может быть разложено без остатка на причинно-следственные звенья. Развитие – целостный процесс.
Регуляции в раннем развитии.
I. Опыты на зиготе, до начала дробления: от яйцеклетки отрывали часть цитоплазмы при центрифугировании, отсасывали микропипеткой или сливали вместе две оплодотворенные яйцеклетки. Регуляция до целого, за исключением яиц насекомых, при разрушении половой плазмы, и если происходило отторжение 1/3, 1/4 части цитоплазмы. Такие результаты были получены на яйцеклетках полихет, моллюсков, иглокожих, асцид. Результаты этих опытов можно трактовать так, что вещества ооплазмы и структуры карт(?) слоя Þ эквипотенциальны, взаимозаменяемы.
II. Опыты по разделению, сращиванию и перетасовке бластомеров дробящихся зародышей. Э. Геккель, В. Ру, 1883-1869 г. Возможность развития целых организмов из изолированных бластомеров. Позже это явление было обнаружено у большинства беспозвоночных и позвоночных животных. Явление «однояйцевых близнецов», Ганс Дриш, Ганс Шлеман, Герличка, Конклин, Отто Шмидт, Зейдель и другие. У человека спонтанное разделение бластомеров на 80 – 1 двойняшки, 6400 – 1 тройняшки, 51200 – четверо. Однояйцевые близнецы возникают при случайном распадении зародыша на бластомеры.
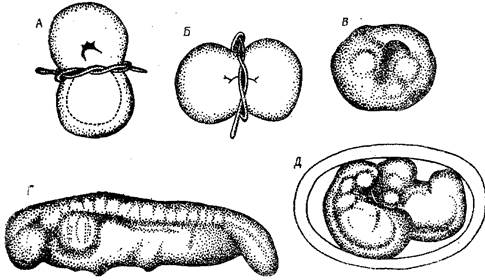
Опыты Г. Шпемана по перешнуровке яиц тритона по фронтальной (А) и сагитальной (Б) плоскостям.
В – формирование недифференцированного комка тканей из вентральной части; Г – развитие полноценного зародыша из дорсальной части, содержащей серый серп; Д – полноценные зародышы, возникшие после сагиттальной перешнуровки.
Искусственная методика на кроликах – Зейдель, 1959 г. Вымывал из яйцеводов ранних зародышей, рассыпал их на бластомеры и имплантировал в матку ® искусственные однояйцевые близнецы с идеальной совместимостью тканей вследствие идентичности геномов.
Мерогония. Е. Гарвей, 1936 г. Опыт по определению носителя генетической информации о развитии, извлечения ядра оплодотворенной ovo путем центрифугирования у морского ежа. Наблюдение: дробление безъядерного зародыша до образования подобия бластулы из комочков цитоплазмы, лишенных ядер. Вывод: за начальный этап онтогенеза – дробление – отвечают носители информации, локализованные в цитоплазме.
Методика изоляции бластомеров. 1. Классический метод Шлемана – перетягивание яиц волосяной петлей, затем она ослабляется и дается возможность проникновению во фрагмент зародыша – одного ядра.
2. Другие способы: растворение яйцевых оболочек или извлечение из среды (для яиц морских беспозвоночных ионов Mg или Са Þ зародыш рассыпается на бластомеры).
Результаты опытов. 1. У гидромедузы Aegineta 1/32 Þ бластомеры тотипотентны, т.е. способны образовывать целое из части.
2. Морские ежи 1/2, 1/4, 1/8 и меридиональные секторы зародыша до стадии бластулы, из перемешанных бластомеров, но при этим нарушается нормальный ход гаструляции – не инвагинация.
У зародышей с преобладанием анимально-вегетативной полярности не получаются пропорциональные зародыши: из анимального полюса – бластулы с ресничками; из вегетативного – преобладание энтодермального материала.