Электрохимический этап мышечного сокращения. 3 страница
Внутренняя среда организма включает в себя организменные жидкости — плазму крови, лимфу, межклеточное вещество и цереброспинальную жидкость. Сохранение стабильности этих жидкостей жизненно важно для организмов, тогда как её отсутствие приводит к повреждению генетического материала.
В отношении любого параметра организмы делятся на конформационные и регуляторные. Регуляторные организмы сохраняют параметр на постоянном уровне, независимо от того, что происходит в среде. Конформационные организмы позволяют окружающей среде определять параметр. Например, теплокровные животные сохраняют постоянную температуру тела, тогда как холоднокровные демонстрируют широкий диапазон температур.
Речь не идёт о том, что конформационные организмы не обладают поведенческими приспособлениями, позволяющими им в некоторой степени регулировать взятый параметр. Рептилии, к примеру, часто сидят на нагретых камнях утром, чтобы повысить температуру тела.
Преимущество гомеостатической регуляции состоит в том, что она позволяет организму функционировать более эффективно. Например, холоднокровные животные, как правило, становятся вялыми при низких температурах, тогда как теплокровные почти так же активны, как и всегда. С другой стороны, регуляция требует энергии. Причина, почему некоторые змеи могут есть только раз в неделю, состоит в том, что они тратят намного меньше энергии для поддержания гомеостаза, чем млекопитающие.
Разные факторы влияют на способность жидкостей организма поддерживать жизнь. В их числе такие параметры, как температура, солёность, кислотность и концентрация питательных веществ — глюкозы, различных ионов, кислорода, и отходов — углекислого газа и мочи. Так как эти параметры влияют на химические реакции, которые сохраняют организм живым, существуют встроенные физиологические механизмы для поддержания их на необходимом уровне.
Гомеостаз нельзя считать причиной процессов этих бессознательных адаптаций. Его следует воспринимать как общую характеристику многих нормальных процессов, действующих совместно, а не как их первопричину. Более того, существует множество биологических явлений, которые не подходят под эту модель — например, анаболизм.
57.Потенциал покоя (ПП) — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ[1]. У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки.
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Источником электромагнитной энергии в любой клетке служит концентрационный элемент, образованный растворами солей, которые неравновесно распределены между цитоплазмой и межклетогной жидкостью, разделенными плазматической мембраной, обладающей неодинаковой проницаемостью для катионов и анионов, на которые диссоциируют эти соли. С другой стороны, сравнение потенциала покоя и равновесного потенциала для конкретного иона позволяет понять и предсказать, куда будет перемещаться этот ион при данном ПП и его изменении (конкретном мембранном потенциале).
Расчеты показывают, что равновесный потенциал при физиологических значениях концентраций ионов по обе стороны плазматической мембраны для важнейших электрогенных ионов в большинстве клеток приблизительно соответствуют.
Функции мембранного потенциала покоя:
1. Поляризация мембраны является условием для возбуждения и торможения.
2.Поляризация определяет объем выделения медиатора из пресинаптического окончания.
3. ПП создает условия для нахождения потенциалзависимых каналов в закрытом состоянии (поляризация мембраны создает условия для формирования потенциала действия).
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 мВ.
Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
Для того чтобы вычислить точное значение мембранного потенциала для модели идеальной клетки, необходимо принять во внимание отдельные ионные токи, протекающие через мембрану. Входящий натриевый ток (iNa) зависит от величины движущей силы для ионов натрия (Vm-ENa), а также от натриевой проводимости мембраны (gNa). Проводимость пропорциональна среднему количеству натриевых каналов, находящихся в открытом состоянии при потенциале покоя: чем больше открытых каналов, тем выше проводимость. Таким образом, натриевый ток равен:

Тот же подход справедлив и для калия и хлора:
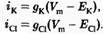
Если принять iCl, = 0 (т.е. ионы хлора находятся в равновесии), то, для сохранения неизменного значения мембранного потенциала, калиевый и натриевый токи должны быть равны по величине и противоположны по направлению:
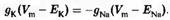
Рассмотрим это уравнение более подробно. Предположим, что gK значительно больше gNa. Тогда, для того чтобы токи были равны, движущая сила для выхода калия должна быть значительно меньше движущей силы для натриевого входа. Другими словами, мембранный потенциал должен быть ближе к ЕK, чем к ENa. Соответственно, если значение gNa велико, то мембранный потенциал будет ближе к ENa.
Преобразовав выражение, получим формулу для мембранного потенциала:

Если по той или иной причине ионы хлора не находятся в равновесии, то формула приобретает несколько более сложный вид:

Вышеприведенные расчеты были первоначально проведены Голдманом) и, независимо от него, Ходжкином и Катцем), с той лишь разницей, что вместо равновесных потенциалов и проводимостей они оперировали ионными концентрациями снаружи ([Na] 0,. .) и внутри ([Na] i,. .) клетки, а также проницаемостью мембраны для каждого из ионов (pNa···):

Так же как и ранее, хлорные компоненты выражения не рассматриваются, если ионы хлора находятся в равновесии. Уравнение называется "уравнением ГХК" по фамилиям авторов, или уравнением постоянного поля, поскольку одним из допущений при выводе уравнения является равномерное распределение градиента напряжения (т.е. "поля") на всем протяжении мембраны.
59. Гладкомышечные эл-ты стенки кровен. сосуда постоянно находятся в состоянии умеренного напряжения – сосудистого тонуса. Существует 3мех-ма рег-ции сос. тонуса: ауторегуляция, нервная регуляция, гуморальная регуляция. Ауторегуляция обеспечивает изменение тонуса гладкомышечных клеток под влиянием местного возбуждения. Значение: поддержание на постоянном уровне объема крови, поступающей к органу (наиболее выражен механизм в почках, печени, легких, головном мозге). Нервная регуляция сосудистого тонуса осуществляется вегетативной нервной системой, которая оказывает сосудосуживающее и сосудорасширяющее действие. Гуморальная регуляция осуществляется веществами системного и местного действия. К веществам системного действия относятся ионы Са, К, Na, гормоны. Ионы Ca вызывают сужение сосудов, ионы K оказывают расширяющее действие. В продолговатом мозге локализуется сосудодвигательный центр, который состоит из двух областей – прессорной и депрессорной. Возбуждение нейронов прессорной области приводит к повышению тонуса сосудов и уменьшению их просвета, возбуждение нейронов депрессорной зоны обусловливает понижение тонуса сосудов и увеличение их просвета. Тонус сосудодвигательного центра зависит от нервных импульсов, постоянно идущих к нему от рецепторов рефлексогенных зон. Особенно важная роль принадлежит аортальной и каротидной рефлексогенным зонам. Барорецепторы дуги аорты и каротидного синуса одинаково влияют на центр продолговатого мозга. Снижение давления в каротидном синусе вызывает спазм мышц сосудов. Стимуляция аортальных телец также вызывает вазоконстрикторный эффект.
58.Согласно функциональной классификации, ионные каналы группируются по способам управления их состоянием на следующие виды: неуправляемые (независимые), потенциал-управляемые (потенциал-чувствительные, потенциал-зависимые), лиганд-управляемые (хемо-управляемые, хемочувствительные, хемозависимые, лиганд-зависимые, рецептор-активируемые), опосредованно-управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер-управляемые, управляемые метаботропными рецепторами). Наиболее часто встречаются два типа каналов: ионные каналы с лиганд-зависимыми воротами (находятся, в частности, в постсинаптической мембране нервно-мышечных соединений) и ионные каналы с потенциал-зависимыми воротами. Лиганд-зависимые каналы превращают химические сигналы, приходящие к клетке, в электрические; они необходимы, в частности, для работы химических синапсов. Потенциал-зависимые каналы нужны для распространения потенциала действия. Потенциал-зависимые ионные каналы: Эти каналы отвечают за распространение потенциала действия, они открываются и закрываются в ответ на изменение мембранного потенциала. Например, натриевые каналы. Если мембранный потенциал поддерживается на уровне потенциала покоя, натриевые каналы закрыты и натриевый ток отсутствует. Если мембранный потенциал сдвигается в положительную сторону, то натриевые каналы откроются, и в клетку начнут входить ионы натрия по градиенту концентрации. Через 0,5 мс после установления нового значения мембранного потенциала, этот натриевый ток достигнет максимума. А еще через несколько миллисекунд падает почти до нуля. Это значит, что каналы через некоторое время закрываются вследствие инактивации, даже если клеточная мембрана остается деполяризованной. Но закрывшись, они отличаются от состояния, в котором находились до открытия, теперь они не могут открываться в ответ на деполяризацию мембраны, то есть они инактивированны. В таком состоянии они останутся до тех пор, пока мембранный потенциал не вернется к исходному значению и не пройдет восстановительный период, занимающий несколько миллисекунд. Для каналов характерна ионная специфичность (каналы одного типа пропускают только ионы калия, другого — только ионы натрия), селективность (это избирательно повышенная проницаемость ионного канала для определённых ионов и пониженная для других), проницаемость (это способность открываться или закрываться при определённых управляющих воздействиях на канал), инактивация, блокировка, пластичность.
61. Чем короче в возбудимых образованиях длительность рефрактерной фазы, тем с большей частотой они могут отвечать на повторные раздражения, и наоборот. Скорость протекания процессов возбуждения была названа Н.Е. Введенским функциональной подвижностью и явл-ся максимально доступное количество циклов возбуждения, воспроизводимых в единицу времени (секунду). Таким образом, зная величину лабильности, можно определить полную длительность одного цикла возбуждения.
Функциональная подвижность непостоянна и может, как увеличиваться, так и уменьшаться. Особенно четко выражена эта изменчивость у малолабильных образований, например у мышцы сердца. В процессе деятельности может происходить сонастраивание, т.е. «усвоение» ритма (А.А. Ухтомский). Органы и ткани, изменяя свою лабильность, в состоянии усваивать как более высокие, так и более низкие ритмы. Усвоение ритма различными системами орг-ма может наблюдаться в процессе врабатывания при занятиях физическими упражнениями. Для каждой ткани характерен некоторый диапазон (в пределах шкалы функциональной подвижности) рабочего ритма, в котором её ф-ия оказывается наиболее эффективной по производительности и энергической экономичности и может продолжаться более длительное время.
62. Мочеиспускание (М.)- сложный рефлекторный акт и осущ-ся сокращением гладкой мускулатуры стенок мочевого пузыря (m. detrusor) и одновр-ым расслаблением обоих сфинктеров - сфинктера мочевого пузыря и сфинктера мочеиспускат. канала.
Мочеиспускание осущ-ся в 3 фазы: открытие, опорожнение и закрытие.
В фазе открытия детрузор сокращ-ся, а поперечнопол. мускулатура уретры и тазового дна расслабл-ся. Уретра, прочно закрепленная в мочеполовой диафрагме, оттягивается в каудальном напр-ии, пластинка основания мочевого пузыря воронкообразно вытягивается. Устья мочеточников при этом закрыв-ся, что предотвращает обратное поступление мочи из мочевого пузыря в мочеточники. Сигналы от высших регулирующих центров НС и мотонейронов II-IV крестцового сегмента спинного мозга, поступающие по половым нервам, вызывают расслабление уретрального сфинктера. Моча при этом может свободно проходить по уретре наружу. На фоне сокращения детрузора мочевой пузырь приобретает шаровидную ф-му. Давление в его просвете повышается. Этому спос-ют напряжение мышц брюшной стенки, тазового дна и умеренное повышение внутрибрюшного давления.
Начало фазы опорожнения сопров-ся незначительным нарастанием, а затем стабилизацией внутрипузырного давления за счет постоянного оттока мочи по уретре. Если раздражение рецепторов уменьш-ся, то М. может прекратиться. Однако при отс-ии нарушений возбудимости детрузора этого не происходит, т.к. поступающая в задний отдел уретры моча рефлекторно спос-ет открытию сфинктера и сокращению детрузора, к-ое подкрепляет раздражение механо- и барорецепторов. Это в еще большей степени усиливает рефлекторное опорожнение мочевого пузыря, открытие шейки и замыкательного аппарата и расслабление уретрального сфинктера.
Чем выше степень растяжения детрузора, тем сильнее его сокращение, полнее открытие шейки мочевого пузыря и больше скорость его опорожнения. В норме она зависит от величины внутрипузырного давления и диаметра внутреннего отверстия мочеиспускательного канала. Опорожнению мочевого пузыря спос-ет давление, создающееся в его просвете (детрузорное и внутрибрюшное). В формир-ии детрузорного давления участвует только гладкая мускулатура мочевого пузыря. Детрузорное давление явл-ся главным мех-мом открытия шейки мочевого пузыря и его опорожнения. Внутрибрюшное давление может произв-но изменяться в больших пределах, поскольку в основе его повышения лежит деят-ть мышц живота, спины и таза. Этот компонент внутрипузырного давления влияет на скорость опорожнения мочевого пузыря лишь при условии открытия его шейки.
В конце М. (фаза закрытия) сокращ-ся поперечнопол. мускулатура уретры и тазового дна, приподнимается шейка мочевого пузыря, сглаживается пластинка его основания, расслабляется детрузор и закрывается внутреннее отверстие уретры. У мужчин выделение остатков мочи происх-т под влиянием сокращения бульбокавернозных мышц и может сопров-ся умеренным повышением внутрипузырного давления. При мочеиспускании, особенно у женщин, на фоне интенсивного поступления мочи по уретре детрузорный компонент внутрипузырного давления может резко уменьш-ся, и эвакуация мочи может происходить почти пассивно под влиянием внутрибрюшного давления. В этом случае сод-ое мочевого пузыря как бы выжимается по уретре наружу. Спиральное направление гладкомышечных волокон детрузора, формир-ие воронки в зоне шейки мочевого пузыря и заднего отдела уретры создают вращательный момент при движении мочи, к-ый спос-т скорейшему опорожнению мочевого пузыря с минимальными энергетическими затратами.
Выделительная ф-ия кожи преимущ-но обеспеч-ся деят-тью потовых желез и, в меньшей степени, сальных желез. В среднем у человека за сутки выд-ся от 300 до 1000 мл пота. Кол-во пота зависит от t окружающей среды и интенс-ти энергет. метаболизма. В условиях большой физ. нагрузки и высокой t воздуха потоотделение может возрастать до 10 л в сутки. Составы пота и плазмы крови отличаются, следовательно, пот явл-ся не простым фильтратом плазмы, а секретом потовых желез. С потом из орг-ма выводится в покое до 1/3 общего кол-ва экскретируемой воды, 5-10% всей мочевины, мочевая к-та, креатин, хлориды, Na, K, Ca, орган. в-ва, липиды, микроэлементы. Ч/з кожу может выдел-ся даже больше Са, чем выводится с мочой. При недостаточности ф-ии почек / печени возрастает выделение ч/з кожу вв, обычно экскретируемых с мочой - мочевины, ацетона, желчных пигментов и др. С потом выдел-ся пепсиноген, амилаза и щелочная фосфатаза, отражая тем самым функцион. сост-ие органов пищеварения. Регуляция потоотд-ия осущ-ся нейрогенно симпатическими холинергическими влияниями, а также гормонами - вазопрессином, алъдостероном, гормонами щитовидной железы и половыми стероидами.
Секрет сальных желез на 2/3 состоит из воды, а 1/3 составляют неомыляемые соединения - холестерин, сквален (алифатический углеводород), аналоги казеина, пр-ты обмена половых гормонов, кортикостероидов, витаминов и ферментов. В выделит. системе сальные железы не имеют большой значимости, т.к. за сутки выд-ся всего лишь около 20 г секрета. Регуляция сальных желез обеспеч-ся в основном половыми и надпочечниковыми стероидами.
Выделительная ф-ия легких. Процессы газообмена, происх-ие в легких, обеспеч-т удаление из внутренней среды орг-ма летучих метаболитов и экзогенных в-в – СО2, аммиака, ацетона, этанола и др. Кроме того, за счет мерцательного эпителия удаляются пр-ты обмена в-в самой легочной ткани и эпителия воздухоносных путей, например, пр-ты деградации сурфактанта. Ч/з слизистую оболочку дыхат. путей испаряется значительное кол-во воды (от 400 мл в покое до 1 л при усиленном дыхании), а при повышении проницаемости аэрогематического барьера из крови могут в избытке выделяться пурины, аденозин- и гуанозинмонофосфаты.
Выделительная ф-ия желудка обесп-т выведение в составе желудочного сока пр-тов метаболизма (мочевины, мочевой кислоты), лекарственных и ядовитых в-в (ртуть, иод, салицилаты, хинин).
Выделительная ф-ия кишечника состоит: Во-первых, в выделении пр-тов распада пищевых в-в, не подвергшихся всасыванию в кровь, и представляющих излишние или вредные для орг-ма соединения. Во-вторых, кишечник экскретирует в-ва, поступившие в его просвет с пищеварительными соками (желудочным, поджелудочным) и желчью. При этом многие из них в кишечнике подвергаются метаболизму и с калом выдел-ся не сами в-ва, а их метаболиты, например, метаболиты билирубина желчи. В-третьих, стенка кишечника способна экскретировать из крови ряд в-в, среди к-ых особое значение имеет экскреция плазменных белков. При чрезмерности этого процесса возникает избыточная потеря орг-мом белка, ведущая к патологии. Из крови кишечный эпителий экскретирует соли тяжелых металлов, магний, почти половину всего выделяемого орг-мом кальция. Вместе с экскрементами выделяется и некоторое количество воды (в среднем около 100 мл/сут).
63. Система органов кровообращения — сердце, сосуды, капилляры — обесп-т непрерывное движение (циркуляцию) крови в орг-ме животных. Кровь приводится в движение сердцем, поэтому изучению ф-ий этого органа придается особое значение. Кровеносные сосуды делятся на артерии, артериолы, капилляры и вены. По артериям кровь течет от сердца к тканям. Артерии по току крови древовидно ветвятся на все более мелкие сосуды и, наконец, превращаются в артериолы, которые в свою очередь распадаются на систему тончайших сосудов — капилляров. Капилляры имеют просвет, почти равный диаметру красных кровяных телец (около 8 мкм). От капилляров начинаются мелкие вены, которые постепенно сливаются и укрупняются. К сердцу кровь притекает по самым крупным венам.
Система кровообращения обесп-т непрерывную циркуляцию крови, необходимую для выполнения кровью транспортных ф-ий: она доставляет к тканям питательные в-ва и кислород и удаляет продукты обмена и углекислый газ. В центре системы кровообращения находится сердце. От него начинается круг кровообращения. Последний делится на две части — большой и малый.
Малый круг кровообращения начинается легочным стволом, который отходит от правого желудочка. По нему кровь доставляется в систему легочных капилляров. От легких кровь оттекает по четырем венам, впадающим в левое предсердие. Здесь заканчивается малый круг кровообращения. Кровь, проходя через капилляры малого круга, отдает углекислый газ и насыщается кислородом. Таким образом, кровь из венозной превращается в артериальную.
Большой круг кровообращения начинается от левого желудочка, из которого кровь поступает в аорту. Из аорты через систему артерий кровь уносится в систему капилляров органов и тканей всего тела. От органов и тканей кровь оттекает по венам и через две полые вены вливается в правое предсердие.
Артерии представляют собой цилиндрической формы трубки, по которым кровь транспортируется от сердца к органам и тканям. Капилляры – это микроскопические (волосные) сосуды, которые находятся в тканях и соединяют артерии с венами. Вены. Работа сердца интересовала ученых еще в глубокой древности, но понять принцип кровообращения долгое время не удавалось. Центром кровообращения признавали печень. Эволюция сердечно-сосудистой системы. Совершенствование ф-ий сердца и сосудов у разных животных явл-ся результатом длительного филогенетического развития. Строение сердца изменялось в процессе филогенеза.
У млекопитающих и птиц сердце разделено на две половины, не сообщающиеся между собой. В сердце плода млекопитающих между предсердиями имеется отверстие, которое к моменту рождения постепенно зарастает. В процессе развития изменилась и питательная жидкость. С появлением гастроваскулярной системы по сосудам течет водянистая жидкость, не имеющая белка и азотсодержащих в-в. Это так называемая гидролимфа. В последующем она заменилась гемолимфой — жидкостью, богатой органическими и неорганическими в-вами. При дальнейшей эволюции появилась кровь с присущими ей ф-иями.
64. Любая приспособительная реакция возможна только в том случае, если для ее возникновения есть побудительные причины. Ими явл-ся различные ф-ры внешней и внутренней среды. В физиологии они получили название раздражитель, стимул, возмущающее воздействие. Процесс воздействия раздражителя на живое образование называется раздражением.По своей энергетической природе раздражители могут быть химическими, электрическими, механическими, температурными, радиационными и др. По биологическому значению - адекватными (от лат. adaequatus - соответствующий, специальный) и неадекватными (общими). Адекватные раздражители - это раздражители, которые при минимальной энергии раздражения вызывают возбуждение в рецепторах и клетках, обладающих специальной способностью реагировать на данный раздражитель. Например, для чувствительных клеток сетчатки глаза адекватным раздражителем явл-ся часть спектра электромагнитных волн (световые излучения), для органов слуха - частота воздушных колебаний в пределах от 16 до 20000 герц, для мышцы - нервный импульс. Все остальные раздражители для перечисленных образований будут неадекватными. Неадекватные, или общие раздражители - это те виды воздействий, которые могут восприниматься многими возбудимыми образованиями и вызывать ответные реакции (как возбуждение, так и торможение) только при относительно значительной силе и длительности своего воздействия. Примером может служить ощущение вспышки света при механическом раздражении сетчатки глаза в результате сдавливания глазного яблока. Следовательно, многие (как адекватные, так и неадекватные) воздействия внешней среды на орг-м и сдвиги во внутренней его среде могут выполнять роль раздражителей, вызывающих ответную реакцию. Однако реакция на адекватный раздражитель возникает легче и имеет более совершенный приспособительный характер, чем реакция на неадекватный раздражитель. Способность живого образования отвечать на действия раздражителя изменениями своих физиологических свойств и возникновением состояния возбуждения называется возбудимостью.
65. Сон — такое состояние орг-ма животного и человека, которое характеризуется прекращением или значительным снижением двигательной активности, понижением ф-ии анализаторов, сокращением контакта с окружающей средой, более или менее полным отключением сознания. Естественный физиологический сон носит периодический характер. Он может быть сезонным (зимняя или летняя спячка у животных) и суточным (ночной или дневной) и у человека, и у животных.
Гипнотический сон представляет собой частичный (парциальный) сон, характеризующийся неполным торможением коры больших полушарий головного мозга при сохранении некоторого контакта человека с внешним миром. Наркотический сон возникает под влиянием действия на орг-м различных в-в естественного и фармакологического происхождения — эфира, хлороформа, закиси азота, многих ядов и т. д. Патологический сон возникает вследствие различных расстройств центральной нервной системы. Физиологическая картина сна. Во время сна наблюдается резкое угнетение тонуса скелетной мускулатуры, гладкая же мускулатура сфинктеров, напротив, находится в состоянии повышенного тонуса. Частота дыхания и сердцебиения заметно урежается, но глубина дыхания увеличивается. При переходе от бодрствования ко сну и при пробуждении иногда наблюдаются фазовые состояния, называемые гипнотическими фазами, во время которых возникают специфические изменения рефлекторной деятельности. Так, если при нормальном бодрствовании сильные раздражители вызывают более сильные ответные реакции, чем слабые, то при фазовых состояниях происходят следующие изменения: I фаза — уравнительная — характеризуется одинаковой величиной (уравниванием) условнорефлекторных реакций на слабые и сильные раздражители; II фаза — парадоксальная — величина рефлексов на сильные раздражители большая, чем на слабые; III фаза — ультрапарадоксальная — наблюдается реже, чем другие фазы, и заключается в том, что на отрицательные условные раздражители, обычно не вызывающие ответной реакции, возникает условная реакция, а на прочно выработанные — не возникает.
Биоэлектрическая активность во время сна и бодрствования. Описание и систематизация результатов регистрации биоэлектрической активности мозга во время сна выявила характерную динамику происходящих в ЭЭГ изменений. На этом основании были выделены различные стадии сна
Первая стадия характеризуется развитием сонливости и легкой дремоты. Вторая стадия возникает по мере углубления сна. Третья стадия характеризуется дальнейшим углублением сна и появлением в ЭЭГ высокоамплитудных (свыше 160 мкВ) потенциалов. Четвертая стадия, соответствующая глубокому сну.
Физиологические теории сна. Вначале XX в. Наиболее широкое распространение получила гуморальная или токсическая теория сна, исходящаа из гипотезы, что в процессе утомления происходит накопление в крови токсических в-в, что и вызывает сон. Согласно теории И.П. Павлова, сон представляет собой разлитое генерализованное торможение, охватывающее кору больших полушарий. X. Мэгун и Г. Моруцци (1956, 1959), раздражая ретикулярную формацию мозга, обнаружили развитие реакции пробуждения у спящего и даже наркотизированного животного, а в ЭЭГ реакцию десинхронизации. Эти опыты показали, что в центральной нервной системе существует две функционально противодействующие системы, регулирующие сон и бодрствование (рис. 144). Электрическое раздражение ретикулярной формации ствола мозга вызывает у сонного животного реакцию пробуждения, а находящегося в состоянии бодрствования — настораживание. И, наоборот, при повреждении этого отдела центральной нервной системы животное впадает в длительный сон, из которого вывести животное практически невозможно. П.К. Анохин объединил разноречивые экспериментальные факты в единую теорию сна и бодрствования, рассматривающую сон как результат определенных циклических изменений во взаимоотношениях коры и важнейших подкорковых образований и, в частности, гипоталамуса и области ретикулярной формации ствола мозга. Бодрствование. Переход от состояния сна к бодрствованию может осуществляться как в результате биохимических сдвигов в орг-ме, так и изменения функциональных отношений между корой, гипоталамусом и ретикулярной формацией либо под влиянием раздражителей из внешней среды. Состояние бодрствования характеризуется повышением анализаторной и двигательной активности и поддерживается в орг-ме на протяжении определенного времени в результате множества ф-ров, воздействующих из внешней и внутренней среды орг-ма.