Структура и функция центрального отдела обонятельного анализатора
Структурно-функциональная организация обонятельного анализатора.
Обонятельную систему человека можно разделить на три отдела: периферический (нейроэпителий, выстилающий верхнюю камеру носа), промежуточный (обонятельная луковица и переднее обонятельное ядро) и центральный (палеокорковая, таламическая, гипоталамическая и неокорковая проекции).
Периферический отдел
Нос человека имеет три камеры: нижнюю, среднюю и верхнюю.
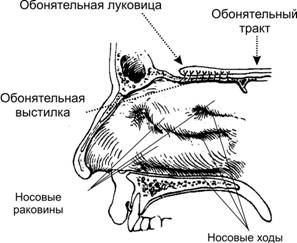
Рис. 1. Топография обонятельной выстилки, обонятельной луковицы и обонятельного тракта
Нижняя и средняя камеры выполняют, по сути, санитарную роль, согревая и очищая вдыхаемый воздух.
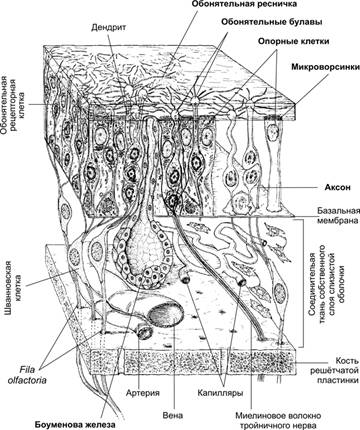
Рис. 2. Орган обоняния [11]. В эпителии обонятельной выстилки расположены обонятельные рецепторные нейроны. Их наружные отростки заканчиваются обонятельными булавами, от которых в разные стороны параллельно поверхности эпителия отходят обонятельные волоски. Аксоны рецепторных нейронов в составе проходят через lamina cribrosa решётчатой кости и образуют обонятельных нитей (fila olfactoria) обонятельного нерва синапсы с нейронами обонятельной луковицы (bulbus olfactorius). Многочисленные опорные клетки окружают рецепторные клетки.
Стенки верхней камеры выстланы нейроэпителием. Обонятельный эпителий, выстилающий обонятельную область носа, имеет толщину 100-150 мкм и содержит рецепторные, а также расположенные между ними опорные клетки. Количество обонятельных рецепторов весьма велико и в значительной степени определяется площадью, занимаемой обонятельным эпителием и плотностью рецепторов в нем. Всего в обонятельной области у человека на площади около 10 см2.содержится приблизительно 107 рецепторов. Число их у других позвоночных может быть и гораздо больше (например, у немецкой овчарки-2,2- 108). Обонятельные клетки, подобно вкусовым, регулярно замещаются; при этом, по-видимому, не все они функционируют одновременно.

Рис. 3. Строение обонятельного эпителия.
Обонятельные рецепторыявляются первичными биполярными сенсорными клетками. Обонятельная рецепторная клетка имеет веретенообразную форму, в виде обонятельной булавы, от которой отходит по два отростка: от верхней части - дендрит, несущий 6-12 ресничек (цилии),а от основания - аксон. Реснички, внутренняя структура которых иная, чем у обычных киноцилий, погружены в слой слизи, вырабатываемой боуменовыми железами. Пахучие вещества, переносимые вдыхаемым воздухом, вступают в контакт с мембраной ресничек - наиболее вероятное место взаимодействия между стимулирующей молекулой и рецептором. Центральные отростки обонятельных рецепторов представляют собой немиелинизированные нервные волокна, которые у млекопитающих собираются в пучки по 10—15 волокон (обонятельные нити — filea olfactoriae) и, пройдя через отверстия решетчатой кости, направляются к обонятельной луковице мозга.
Слизистая оболочка носа, кроме того, содержит свободные окончания тройничного нерва, часть которых также способна реагировать на запахи. Эти ноцирецепторы стимулируют раздражающие вещества, и этот раздражающий компонент — часть характеристики «запаха» ряда веществ (аммиака, хлорной извести и др.). Другими словами, эти болевые нервные окончания ответственны за чихание, слёзы, задержку дыхания и другие рефлексы, вызванные раздражением слизистой оболочки носа.
В области глотки обонятельные стимулы способны возбуждать волокна языкогло-точного (IX) и блуждающего (X) нервов.
Восприятие запахов. Последовательность событий. Молекулы пахучего вещества взаимодействуют со специализированными белками, встроенными в рецептор нейросенсорных клеток. При этом происходит адсорбция раздражителей на хеморецепторной мембране. Согласно стереохимической теории, этот контакт возможен в том случае, если форма молекулы пахучего вещества соответствует форме рецепторного белка в мембране («ключ — замок»). Слизь, покрывающая поверхность хеморецептора, является структурированным матриксом. Слизь содержит воду, гликозаминогликаны, АТ, связывающие молекулы одорантов белки, ферменты и полностью обновляется в течение 10 мин. Она контролирует доступность рецепторной поверхности для молекул раздражителя и способна изменять условия рецепции. Слой слизи, покрывающий обонятельный эпителий и предохраняющий его от высыхания, постоянно возобновляется благодаря секреции и движению киноцилий окружающего эпителия.
Современная теория обонятельной рецепции предполагает, что начальным звеном этого процесса могут быть два вида взаимодействия: первое — контактный перенос заряда при соударении молекул пахучего вещества с рецептивным участком, и второе — образование молекулярных комплексов и комплексов с переносом заряда. Эти комплексы обязательно образуются с белковыми молекулами рецепторной мембраны, активные участки которых выполняют функции доноров и акцепторов электронов. Существенным моментом этой теории является положение о многоточечных взаимодействиях молекул пахучих веществ и рецептивных участков. Вслед за этим взаимодействием изменяется форма белковой молекулы, обонятельный белок активирует, как и в случае фоторецепции, ГТФ - связывающий белок (G-белок), а тот в свою очередь- фермент аденилатциклазу, синтезирующую цАМФ. Активизируются натриевые каналы, происходит деполяризация мембраны рецептора и генерируется рецепторный потенциал, который, достигнув критической величины, обеспечивает возникновение ПД в аксонном холмике нейросенсорной клетки.
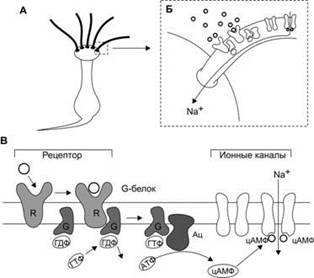
Рис. 5. Трансформация сигнала в обонятельном рецепторе. А — рецепторный обонятельный нейрон; Б — вход Na+ в клетку; В — пахучие молекулы соединяются с рецептором (R). Рецептор активирует Gбелок (G), Gбелок активирует аденилатциклазу (Ац), образующийся цАМФ открывает Na+каналы.
Система инозитолтрифосфата (ИТФ) также имеет отношение к механизму хемовосприятия в органе обоняния. При действии некоторых пахучих веществ быстро возрастает уровень ИТФ, который взаимодействует с Ca2+ каналом в плазмолемме обонятельных рецепторных нейронов.
Таким образом, системы вторых посредников цАМФ и ИТФ взаимодействуют между собой, обеспечивая лучшее различение запахов.
Суммарный электрический потенциал, регистрируемый от поверхности обонятельного эпителия, называется электроольфактограммой.
Особенности кодирования обонятельной информации. Отдельная нейросенсорная клетка способна реагировать на значительное число различных пахучих веществ. В связи с этим различные обонятельные рецепторы (так же, как и вкусовые) имеют перекрывающиеся профили ответов. Каждое пахучее вещество дает специфическую картину возбуждения в популяции чувствительных клеток, при этом уровень возбуждения зависит от концентрации вещества.
У человека обоняние очень чувствительно, хотя и известно, что у некоторых животных этот аппарат более совершенен. При действии очень малых концентраций веществ ощущение неспецифично; в несколько более высоких концентрациях запах не только выявляется, но и идентифицируется. Например, запах скатола в низких концентрациях не так неприятен, тогда как при превышении некоего порога проявляется типичный для этого вещества отталкивающий запах. Таким образом, необходимо различать порог выявлениязапаха и порог его распознавания.
В волокнах обонятельного нерва при электрофизиологическом исследовании обнаружена непрерывная импульсация, обусловленная подпороговым воздействием пахучих веществ. При пороговой и сверхпороговой концентрациях различных пахучих веществ возникают разные типы (паттерны) электрических импульсов, которые приходят одновременно в различные участки обонятельной луковицы. При этом в обонятельной луковице создается своеобразная мозаика из возбужденных и невозбужденных участков. Предполагают, что это лежит в основе кодирования информации о специфичности запахов
Промежуточный отдел. Обонятельная луковица.Гистологически обонятельная луковица подразделяется на несколько слоев, характеризующихся клетками специфической формы, снабженными отростками определенного типа с типичными видами связей между ними. Основными чертами обработки информации в обонятельной луковице являются: 1) заметная конвергенциячувствительных клеток на митральных клетках, 2) выраженные тормозные механизмы и 3) эфферентный контрольимпульсации, входящей в луковицу. В клубочковом (гломерулярном) слое аксоны приблизительно 1000 обонятельных клеток оканчиваются на первичных дендритах одной митральной клетки.Эти дендриты также образуют реципрокные дендродендритные синапсы с перигломерулярными клетками.Контакты между митральными и перигломерулярными клетками - возбуждающие, а противоположного направления - тормозные.
Проводниковый отдел обонятельного анализатора начинается нейросенсорными клетками (первые нейроны), аксоны которых, проходя в полость черепа через отверстие в решетчатой кости, контактируют с крупными митральными клетками обонятельных луковиц, представляющими второй нейрон. Эти клетки имеют главный дендрит, дистальные веточки которого образуют с аксонами нейросенсорных обонятельных клеток синапсы, называемые гломерулами. Центральные отростки клеток обонятельных луковиц в составе обонятельного тракта направляются к обонятельному треугольнику, переднему продырявленному веществу и прозрачной перегородке, где и заканчиваются(третьи нейроны). Аксоны третьих нейронов заканчиваются в крючке гиппокампа, который является корковым концом обонятельного анализатора.
Однако одни авторы считают, что волокна обонятельного тракта отдельными пучками идут прямо в кору большого мозга, минуя зрительные бугры. Другие авторы считают, что отростки второго нейрона могут направляться и в передние ядра зрительного бугра.
Структура и функция центрального отдела обонятельного анализатора
В составе обонятельного мозга различают медиальную и латеральную обонятельные области. Аксоны митральных клеток образуют латеральный обонятельныйтракт. Волокна латерального обонятельного тракта оканчиваются в различных отделах переднего мозга: в переднем обонятельном ядре, латеральной части обонятельного бугорка, препириформной и периамигдалярной областях коры, а также в прилегающей к ней кортико-медиальной части миндалевидного комплекса, включая ядро латерального обонятельного тракта, в которое, как полагают, приходят также волокна из добавочной обонятельной луковицы. Синапсы с нейронами высших порядков обеспечивают связь с гипокампом,ачерез амигдалярный комплекс - с вегетативными ядрамигипоталамуса. Связи обонятельной луковицы с гиппокампом, энторинальной областью пириформной коры и другими отделами обонятельного мозга у млекопитающих осуществляются через одно или несколько переключений.
Нейроны медиальной обонятельной области (ядра перегородки) проецируются в гипоталамус и другие области, контролирующие поведение. От первичной обонятельной коры нервные волокна направляются к медиовентральному ядру таламуса, к которому имеется также прямой вход от вкусовой системы. Волокна медиовентрального ядра таламуса, в свою очередь, направляются к фронтальной области новой коры, которая рассматривается как высший интегративный центр обонятельной системы. Волокна от препириформной коры и обонятельного бугорка идут в каудальном направлении, входя в состав медиального пучка переднего мозга. Окончания волокон этого пучка обнаруживаются в латеральном преоптическом ядре, в латеральной области гипоталамуса. Из описания этих связей становится понятной тесная связь обоняния с пищевым и половым поведением млекопитающих. Эксперименты на животных также показали, что реакции нейронов обонятельного тракта могут быть изменены инъекцией тестостерона. Таким образом, возбуждение обонятельных нейронов находится под влиянием половых гормонов.
Нейроны, отвечающие на обонятельные стимулы, обнаружены также в ретикулярной формации среднего мозга. Связь с лимбической системой объясняет присутствие эмоционального компонента в обонятельном восприятии. Запах может вызывать ощущение удовольствия или отвращения (гедонические компоненты ощущения), при этом меняется состояние организма.
Классификация пахучих веществ и запахов.Человек способен различать запах нескольких тысяч различных веществ. Первая группа пахучих веществ — ольфактивные вещества, которые раздражают только обонятельные клетки. К ним относятся запах гвоздики, лаванды, аниса, бензола, ксилола. Вторая группа — такие вещества, которые одновременно с обонятельными клетками раздражают свободные окончания тройничных нервов в слизистой оболочке носа.К ним относятся запах камфоры, эфира, хлороформа
На основании некоторых психофизиологических наблюдений восприятия запахов человеком выделяют 7 первичных запахов: мускусный, камфарный, цветочный, эфирный, мятный, острый и гнилостный.
Единой и общепринятой классификации запахов не существует. Мы не можем охарактеризовать запах, не называя вещества или предмета, которому они свойственны. Так, мы говорим о запахе камфоры, роз, лука, в некоторых случаях обобщаем запахи родственных веществ или предметов, например цветочный, фруктовый запах и др. Считают, что возникающее многообразие различных запахов является результатом смешения «первичных запахов».
На остроту обоняния влияют многие факторы, например голод, который повышает остроту обоняния; беременность, когда возможно не только обострение обонятельной чувствительности, но и ее извращение.
Особенности адаптации обонятельного анализатора. Адаптация к действию пахучего вещества в обонятельном анализаторе согласно одних авторов происходит сравнительно медленно (в течение десятка секунд или минут, других достаточно быстро (50% в течение 1 с).). Она зависит от скорости потока воздуха над обонятельным эпителием и концентрации пахучего вещества. Обычно адаптация проявляется по отношению к одному запаху.
Различают следующие нарушения обоняния:
1) аносмия — отсутствие обонятельной чувствительности.
При этом заболевании порог для определенных пахучих веществ повышается; по крайней мере, в некоторых случаях это обусловлено генетически. Вредные температурные и химические воздействия в зависимости от их природы и типа действия могут вызывать обратимую или необратимую острую или хроническую аносмию или гипосмию.
2) гипосмия — понижение обоняния. Эти ощущения, не связанные с обонятельным нервом, сохраняются и при нарушениях функции обонятельного эпителия в результате, например, инфекции (гриппа), опухолей (и связанных с этим операций на мозге) или черепно-мозговых травм. При гипофизарном гипогонадизме (синдроме Кальмана) обоняние осуществляется за счет только этих черепно-мозговых нервов, поскольку при этом врожденном заболевании наблюдается аплазия обонятельных луковиц.
3) гиперосмия — повышение,
4) паросмия — неправильное восприятие запахов,
5) нарушение дифференцировки,
6) обонятельные галлюцинации, когда возникают обонятельные ощущения при отсутствии пахучих веществ, или паросмия - неправильное восприятие запаха. Обонятельные галлюцинации неприятного характера (какосмия) проявляются главным образом при шизофрении.
7) обонятельная агнозия, когда человек ощущает запах, но его не узнает. С возрастом в связи с преобладанием инволютивных процессов наблюдаются в основном снижение обонятельной чувствительности, а также другие виды функциональных расстройств обоняния.
Обоняние, половое поведение и память. Между запахом и половой функцией у многих видов животных существует тесная взаимосвязь (скорее всего, реализуемая через cошниково–носовой [вомероназальный] орган Якобсона — участок слизистой оболочки носовых ходов, аналогичный обонятельной выстилке), а использование духов даёт достаточные основания считать, что подобная взаимосвязь существует и у людей (орган Якобсона у человека отсутствует). Обоняние у женщин развито сильнее, и оно обостряется ещё больше в период овуляции. Запах и (в меньшей степени) вкус обладают уникальной способностью оживлять воспоминания, заложенные в долговременной памяти. Этот факт отмечен писателями и экспериментально доказан психологами.
Обонятельная луковица
В обонятельной луковице (рис. 12–1 и 12–4) аксоны рецепторных клеток образуют синапсы с дендритами митральных и пучковых клеток, формируя характерные комплексы— обонятельные клубочки. В каждый клубочек входит (конвергирует) в среднем 25 тыс. аксонов рецепторных клеток, но не любых, а только тех, что имеют идентичные обонятельные рецепторы. В каждом обонятельном клубочке с аксонами рецепторных клеток контактируют дендриты примерно 25 митральных и 60 пучковых клеток. В следующем слое обонятельной луковицы дендриты митральных, зернистых и пучковых клеток образуют реципрокные синапсы. Эти синаптические связи осуществляют контроль за исходящей из обонятельной луковицы информацией, вероятно, закодированной в спектрах ПД.

Рис. 12–4. Архитектоника нейронов и связей в обонятельной луковице. ПК — пучковая клетка, МК — митральная клетка с возвратными коллатералями (К), ЗК — зернистая клетка, ВПЯ — нисходящие в обонятельную луковицу нервные волокна из переднего обонятельного ядра, ВПС — комиссуральные нервные волокна передней спайки мозолистого тела (commissura anterior). Прерывистые линии — границы между анатомическими структурами.
В обонятельную луковицу поступает также импульсация от контралатеральной обонятельной луковицы и нисходящая импульсация из отделов мозга, имеющих отношение к обонянию. Так, к обонятельной луковице идут сигналы из ипси- и контралатеральных передних обонятельных ядер. Стимуляция этих ядер уменьшает электрическую активность обонятельной луковицы.