Переваривание эфиров холестерола
В составе пищи холестерол находится в основном в виде эфиров. Гидролиз эфиров холестерола происходит под действием холестеролэстеразы - фермента, который также синтезируется в поджелудочной железе и секретируется в кишечник. Продукты гидролиза (холестерол и жирные кислоты) всасываются в составе смешанных мицелл.
Всасывание продуктов гидролиза липидов в тонком кишечнике.
Продукты гидролиза липидов – жк с длинным углеводородным радикалом, 2-моноацилглицеролы, холестерол, а также соли жёлчных кислот образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены т.о, что гидрофобные части молекул обращены внутрь мицеллы, а гидрофильные - наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкой кишки. Стабильность мицелл обеспечивается в основном солями жёлчных кислот. Мицеллы сближаются со щёточной каймой клеток слизистой оболочки тонкого кишечника, и липидные компоненты мицелл диффундируют через мембраны внутрь клеток. Вместе с продуктами гидролиза липидов всасываются жирорастворимые витамины A, D, Е, К и соли жёлчных кислот. Жёлчные кислоты далее попадают через воротную вену в печень, из печени вновь секретируются в жёлчный пузырь и далее опять участвуют в эмульгировании жиров. Каждая молекула жёлчных кислот за сутки проходит 5- 8 циклов, и около 5% жёлчных кислот выделяется с калом.
Всасывание жирных кислот со средней длиной цепи, образующихся, например, при переваривании липидов молока, происходит без участия смешанных мицелл. Эти жирные кислоты из клеток слизистой оболочки тонкого кишечника попадают в кровь, связываются с белком альбумином и транспортируются в печень.
Ресинтез таг в кишечнике.После всасывания продуктов гидролиза жиров жирные кислоты и 2-моноацилглицеролы в клетках слизистой оболочки тонкого кишечника включаются в процесс ресинтеза с образованием триацилглицеролов. Жирные кислоты вступают в реакцию этерификации только в активной форме в виде производных коэнзима А, поэтому первая стадия ресинтеза жиров - реакция активации жирной кислоты:
HSКоА + RCOOH + АТФ → R-CO ~ КоА + АМФ + Н4Р2О7.
Реакция катализируется ферментом ацил-КоА-синтетазой. Затем ацил~КоА участвует в реакции этерификации 2-моноацилглицерола с образованием сначала диацилглицерола, а затем триацилглицерола. Реакции ресинтеза жиров катализируют ацилтранеферазы.
Транспорт жиров.Липиды в крови нерастворимы, поэтому для транспорта липидов кровью в организме образуются комплексы липидов с белками - липопротеины. Все типы липопротеинов имеют сходное строение - гидрофобное ядро и гидрофильный слой на поверхности образованный белками, которые называют апопротеинами, и амфифильными молекулами липидов- фосфолипидами и холестеролом. Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина,в котором находятся транспортируемые липиды.
Апопротеины выполняют несколько функций: формируют структуру липопротеинов; взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов; служат ферментами или активаторами ферментов, действующих на липопротеины.
Образование хиломикронов.
Жиры, образовавшиеся в результате ресинтеза в клетках слизистой оболочки кишечника, упаковываются в ХМ. Основной апопротеин в составе ХМ - белок апоВ-48. Этот белок закодирован в том же гене, что и белок ЛПОНП - В-100. В кишечнике в результате посттранскрипционных превращений "считывается" последовательность мРНК, которая кодирует только 48% от длины белка В-100, поэтому этот белок называется апоВ-48. Белок апоВ-48 синтезируется в шероховатом ЭР и там же гликозилируется. Затем в аппарате Гольджи происходит формирование ХМ, называемых "незрелыми". По механизму экзоцитоза они выделяются в хилус, образующийся в лимфатической системе кишечных ворсинок, и через главный грудной лимфатический проток попадают в кровь. В лимфе и крови с ЛПВП на ХМ переносятся апопротеины Е и апоС-П); ХМ превращаются в "зрелые". ХМ транспортируют жир к различным тканям, где он утилизируется, поэтому концентрация ХМ в крови постепенно снижается, и исчезают из крови в течение нескольких часов.
Липопротеинлипаза. В крови триацилглицеролы, входящие в состав зрелых ХМ, гидролизуются ферментом липопротеинлипазой. ЛП-липаза связана с гепаринсульфатом, находящимся на поверхности эндотелиальных клеток, выстилающих стенки капилляров кровеносных сосудов. ЛП-липаза гидролизует молекулы жиров до глицерола и 3 молекул жирных кислот. На поверхности ХМ различают 2 фактора, необходимых для активности ЛП-липазы - апоС-П и фосфолипиды. АпоС-П активирует этот фермент, а фосфолипиды участвуют в, связывании фермента с поверхностью ХМ.
ЛП-липаза синтезируется в клетках многих тканей: жировой, мышечной, в лёгких, селезёнке, клетках лактирующей молочной железы.
Нарушения переваривания липидов в желудочно-кишечном тракте: 1. Нарушение секреции желчи. При этом липиды не эмульгируются и поэтому хуже перевариваются и всасываются, из-за чего возникает стеаторея.
2. Нарушение секреции сока поджелудочной железы, что также приводит к стеаторее.
Стеаторея – Повышенной содержание жиров в кале (выше 5 %). При стеаторее нарушается всасывание жирорастворимых витаминов и незаменимых жирных кислот, также нарушается переваривание веществ нелепиднйо природы, поскольку жир обволакивает эти вещества, не давая ферментам расщепить их.
Липопротеины. Все типы липопротеинов имеют сходное строение - гидрофобное ядро и гидрофильный слой на поверхности. Гидрофильный слой образован белками, которые называют апопротеинами, и амфифильными молекулами липидов- фосфолипидами и холестеролом. Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды.В организме синтезируются следующие типы липопротеинов: хиломикроны, (ЛПОНП), ЛППП), ЛПНП) и ЛПВП) . Апопротеины выполняют несколько функций: Формируют структуру липопротеинов; взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов; служат ферментами или активаторами ферментов, действующих на липопротеины.
Основные апопротеины и их функции:
В-48 - основной белок ХМ;
В-100 - основной белок ЛПОНП, ЛПНП, ЛППП, взаимодействует с рецепторами ЛПНП;
С-II - активатор ЛП-липазы, переносится с ЛПВП на ХМ и ЛПОНП в крови;
Е - взаимодействует с рецепторами ЛПНП;
A-I - активатор фермента ЛХАТ.
Функции ЛП плазмы.
| Тип лп | хм | лпонп | лппп | лпнп | лпвп | ||||
| фкнкции | Транспорт липидов из клеток кишечника | Транспорт липидов, синтезируемых в печени | Промежуточная форма превращения ЛПОНП в ЛПНП под действием фермента ЛП-липазы | Транспорт холестерола в ткани | Удаление избытка холестерола из клеток и других липопротеинов. Донор апопротеиновА, С-П | ||||
| Место образования | Эпителий тонкого кишечника | Клетки печени | Кровь | Кровь (из ЛПОНП и ЛППП) | Клетки печени - ЛПВП-пред-шественники | ||||
Дислипопротеинемии- нарушения обмена ЛП крови и, соответственно, нарушения обмена липидов, транспортируемых ЛП. Дислипопротеинемии проявляются чаще всего повышением концентрации либо одного типа ЛП, либо сочетанным увеличением содержания нескольких типов ЛП. Наиболее распространены нарушения обмена холестерола и триацилглицеролов. Нарушения обмена холестерола чаще всего приводят к гиперхолестеролемиии последующему развитию атеросклероза. При атеросклерозе происходит образование на стенках артерий так называемых атеросклеротических бляшек, представляющих собой в основном отложения холестерола. Атеросклеротические бляшки разрушают клетки эндотелия сосудов, и в таких местах часто образуются тромбы. Одна из основных причин развития атеросклероза - нарушение баланса между поступлением холестерола с пищей, его синтезом и выведением из организма. Выведение холестерола ограничено, не превышает 1,2-1,5 г/сут, а поступление с пищей при неправильном питании может превысить этот барьер, поэтому с возрастом постепенно происходит накопление холестерола в организме. Важным фактором развития атеросклероза являются генетические дефекты белков и ферментов, участвующих в обмене холестерола.
| название | Генетический дефект | Изменение липидного обмена |
| Тип I (наследственная недостаточность ЛП-липазы) | Дефект структуры ЛП-липазы Дефект структуры апоС-П | ↑в крови ХМ и ЛПОНП, нет риска атеросклероза, гипертриглицеролемия |
| Тип II (семейная гиперхолестеролемия) | Дефект рецепторов ЛПНП или мутация гена апоВ-100 | ↑концентрацииЛПНП, гиперхолестеролемия, ранний атеросклероз, ксантоматоз |
| Тип III (семейная комбинированная гиперлипидемия, нарушение удаления остаточных липопротеинов из крови) | Дефект в структуре апоЕ, синтез изоформы апоЕ2, которая не взаимодействует с рецепторами | ↑концентрации остаточных ХМ, ЛПОНП, ЛППП, ЛПНП Гиперхолестеролемия, гипертриглицеролемия, ранний атеросклероз, ксантоматоз |
| Типы IV и V (семейна ягипертриглицеролемия) | Генетически гетерогенная группа заболеваний. Избыточная продукция ЛПОНП как результат гиперинсулинемии | ↑концентрацииЛПОНП, ЛПНП,гипертриглицеролемия, умеренная гиперхолестеролемия Атеросклероз, снижение толерантности к глюкозе, ксантоматоз |
Депонирование и МОБИЛИЗАЦИЯ ЖИРОВ
Адипоциты (место депонирования жиров) располагаются в основном под кожей, образуя подкожный жировой слой, и в брюшной полости, образуя большой и малый сальники. Мобилизация жиров, т.е. гидролиз до глицерола и жирных кислот, происходит в постабсорбтивный период, при голодании и активной физической работе. Гидролиз внутриклеточного жира осуществляется под действием фермента гормончувствительной липазы - ТАГ-липазы. Глицерол как водорастворимое вещество транспортируется кровью в свободном виде, а жирные кислоты в комплексе с белком плазмы - альбумином.
Гидролиз триацилглицеролов до жирных кислот и глицерола катализируют три липазы. Первой действует триацилглицероллипаза, затем диацилглицероллипазаи последней -моноацилглицероллипаза. Эти липазы локализованы в разных органоидах. Указанные липазы различаются не только по специфичности, но и по оптимуму рН и температуре действия.
Мобилизация депонированных жиров стимулируется глюкагоном и адреналином. В постабсорбтивный период и при голодании глюкагон, действуя на адипоциты через аденилатциклазную систему, активирует протеинкиназу А, которая фосфорилирует и, таким образом, активирует гормончувствительную липазу, что инициирует липолиз и выделение жирных кислот и глицерина в кровь. При физической активности увеличивается секреция адреналина, который действует через β-адренергические рецепторы адипоцитов, активирующие аденилатциклазную систему . В настоящее время обнаружено 3 типа β-рецепторов: β1, β2, β3, активация которых приводит к липолитическому действию. К наибольшему липолитическому действию приводит активация β3-рецепторов. Адреналин одновременно действует и на α2-рецепторы адипоцитов, связанные с ингибирующим G-белком, что инактивирует аденилатциклазную систему. Вероятно, действие адреналина двояко: при низких концентрациях в крови преобладает его антилиполитическое действие через α2-рецепторы, а при высокой - преобладает липолитическое действие через β-рецепторы.
Для мышц, сердца, почек, печени при голодании или физической работе жирные кислоты становятся важным источником энергии. Печень перерабатывает часть жирных кислот в кетоновые тела, используемые мозгом, нервной тканью и некоторыми другими тканями как источники энергии.
В результате мобилизации жиров концентрация жирных кислот в крови увеличивается приблизительно в 2 раза, однако абсолютная концентрация жирных кислот в крови невелика даже в этот период. Когда постабсорбтивный период сменяется аборбтивным, инсулин активирует специфическую фосфатазу, которая дефосфорилирует гормончувствительную липазу, и распад жиров останавливается.
Распад жирных кислот. 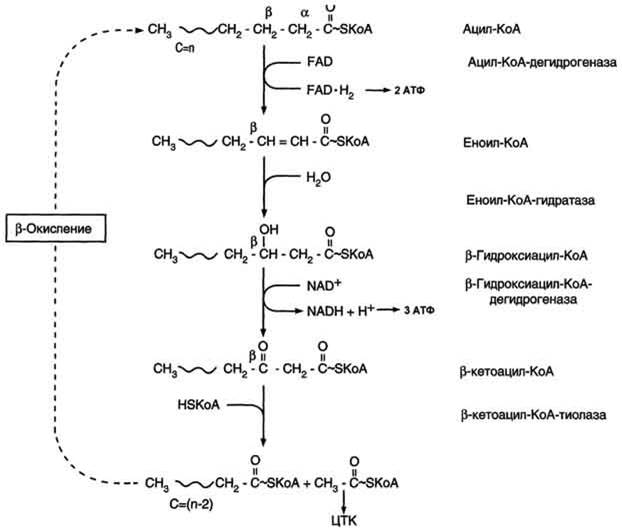
Пояснение: у нас есть ВЖК с количеством углеродов в цепи n. В результате этих 4 реакций у нас получается ацетил-КоА и эта же самая ВЖК, только с количеством углеродов в цепи (n-2) (2 атома ушло на ацетил-КоА). Эта кислота снова вступает в цикл. Таким образом ВЖК будет снова и снова вступать в цикл, пока полностью не расщепиться.
Биологическое значение: образуется много энергии и Ацетил-КоА, который может идти на синтез многих соединений или опять же на ту же энергию.
Локализация в клетке: митохондрии
Энергетический эффект: напрямую зависит от ВЖК. из пальмитиновой кислоты при бета окислении образуется 131 АТФ.