Законы проведения возбуждения по нервному волокну
1. Закон анатомической и физиологической целостности. Как перерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна (блокада натриевых каналов мембраны местными анестетиками, резкое охлаждение и т.п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна ионами калия, накапливающимися при ишемии в межклеточных щелях. Механическая травма, сдавление нерва при воспалительном отеке тканей также могут сопровождаться нарушением функции проведения.
2. Закон изолированного проведения. Нервные стволы состоят из тысяч двигательных, чувствительных и вегетативных волокон, которые иннервируют разные, иногда далеко отстоящие друг от друга и разнородные по структуре и функциям клетки и ткани. Поэтому особенное значение приобретает тот факт, что возбуждение, проходящее по одному нервному волокну, не может переходить на другое волокно. Это обусловлено наличием миелиновой оболочки и жидкости, которая заполняет межклеточные щели. Они препятствуют перескакиванию биотока с возбужденной мембраны на соседние волокна.
3. Закон двустороннего проведения возбуждения. При раздражении среднего участка нервного волокна возбуждение распространяется по нему в обоих направлениях (что можно зафиксировать при помощи электродов). Однако в рефлекторной дуге возбуждение всегда распространяется в одном направлении – от рецепторов к эффекторам. Этот принцип распространения возбуждения в нейронных цепях определяется физиологическими свойствами особого нервного образования – синапса.
Синапсом называется структурное образование, обеспечивающее переход возбуждения с нервного волокна на иннервируемую им клетку (нервную, мышечную или железистую).
Синапс состоит из трех элементов:
– пресинаптическая мембрана, расположенная на синаптической бляшке терминального отдела аксона;
– синаптическая щель, заполненная гликокалексом и межтканевой жидкостью;
– постсинаптическая мембрана, расположенная на иннервируемой клетке. На ней имеются хеморецепторы, чувствительные к определенному виду медиатора. Постинаптическая мембрана также имеет ферменты, разрушающие медиатор после его взаимодействия с хеморецептором.
Медиатор – биологически активное вещество, выделяемое нервным окончанием и осуществляющее передачу возбуждения в химических синапсах. Медиатор синтезируется в теле нейрона и, благодаря механизмам аксонального транспорта, поступает в синаптическую бляшку. В синаптической бляшке медиатор находится в везикулах в строго определенном количестве.
Когда нервный импульс достигает пресинаптической мембраны, в ней активизируются кальциевые каналы. Са++ входит в синаптическую бляшку и способствует прилипанию везикул с медиатором к пресинаптической мембране. Везикулы лопаются и медиатор выходит в синаптическую щель. Выделение медиатора происходит определенными дозами (квантами) и пропорционально количеству нервных импульсов, приходящих к пресинаптической мембране. Выделившийся медиатор диффундирует к постсинаптической мембране, где взаимодействует с ее хеморецепторами (по принципу «ключ к замку»). Постсинаптическая мембрана отвечает изменением проницаемости для ионов Na+ и К+, что приводит к развитию местной деполяризации постсинаптического потенциала (ПСП). По мере высвобождения медиатора деполяризация достигает критического уровня, и возникает ПД (ВПСП), который затем уже механизмом круговых токов, распространяется по иннервируемой структуре.
В тормозных синапсах ионные каналы открываются для ионов К+ и Cl– , в результате чего развивается процесс гиперполяризации постсинаптической мембраны - тормозной постсинаптический потенциал (ТПСП). Но выделение медиатора может происходить не только под действием нервного импульса, но и в покое спонтанно, небольшими порциями. При этом образуются миниатюрные потенциалы, которые повышают возбудимость постсинаптической мембраны и участвуют в обменных процессах иннервируемой клетки.
Синапсы центральной нервной системы имеют следующие основные свойства:
1. Односторонняя передача возбуждения – от пресинаптической мембраны к постсинаптической. Это обусловлено особенностью строения и тем, что постсинаптическая мембрана нечувствительна к электрическим импульсам.
2. Специфичность синапсов – хеморецепторы чувствительны только к определенному виду медиатора.
3. Синаптическая задержка проведения возбуждения (основная составляющая времени рефлекса) – замедление скорости распространения возбуждения в синапсе, в связи с длительностью процессов выделения медиатора из пресинаптического окончания, диффузии его по синаптической щели и процесса взаимодействия его с постинаптической мембраной. Скорость этих процессов в 10 и более раз меньше, чем скорость распространения возбуждения по нерву и в химических синапсах равно 0,2-0,5 м/с.
4. Низкая лабильность – обусловлена синаптической задержкой и высокой утомляемостью. Лабильность синапса составляет 50-100 имп/с.
5. Высокая утомляемость – результат высокого обмена веществ и истощаемости запасов медиатора при продолжительной стимуляции нервного окончания.
Тема 2. Общая физиология центральной нервной системы.
Вопросы для самоподготовки.
I. Рефлекс и рефлекторная дуга.
1. Рефлекс (определение, классификация, биологическая роль).
2. Рефлекторная дуга (строение, локализация и функции составных частей).
3. Время рефлекса (определение; факторы, определяющие его продолжительность; роль центральной задержки)
II. Торможение в центральной нервной системе.
1.Торможение (определение, история открытия, эволюция и онтогенез, функции).
2. Общие и отличительные черты процессов возбуждения и торможения.
3. Виды центрального торможения .
3.1. Пресинаптическое торможение (механизм возникновения торможения, эффекты).
3.2. Постсинаптическое торможение (механизм возникновения торможения, эффекты).
2.3. Торможение в нейронных целях (реципрокное, возвратное).
III. Процессы интеграции и координации в ЦНС.
1. Интегративная деятельность ЦНС, ее уровни (рецепторный, нейрональный, центральный и межцентральный).
2. Координация (определение, факторы ее определяющие).
3. Принципы координации (конвергенция, дивергенция, обратные связи, их характеристика.
4. Эффекты ритмической стимуляции в нейронных цепях.
4.1. Суммация допороговых ритмических стимулов (последовательная, пространственная).
4.2.Окклюзия сверхпороговых стимулов (последовательная, пространственная).
5. Взаимодействие рефлексов (общий конечный путь, иррадиация, доминанта).
Основополагающий материал к теме.
Человеческий организм представляет собой живую систему, которая функционирует по высочайшему принципу саморегулирования. Механизмы саморегуляции организованы иерархически, как, в общем, и уровни организации всех живых систем.
Однако выделяют местную (клеточную, тканевую, органную) и центральную (нервную и гуморальную) виды регуляции. В данном пособии мы не будем рассматривать механизмы этих видов регуляции. Остановимся лишь на том, что главным регуляторным механизмом в организме высших животных и человека является нервная система, главным принципом ее деятельности – рефлекс.
Рефлексом называют любую ответную реакцию организма, осуществляющуюся с участием центральной нервной системы. Морфологической основой рефлекса является рефлекторная дуга (рис. 5), а время, необходимое для проведения возбуждения от рецептора до эффектора называется временем рефлекса. Его продолжительность зависит от количества вставочных нейронов – синаптической (центральной) задержки (см. «Анатомия ЦНС, тема 1).
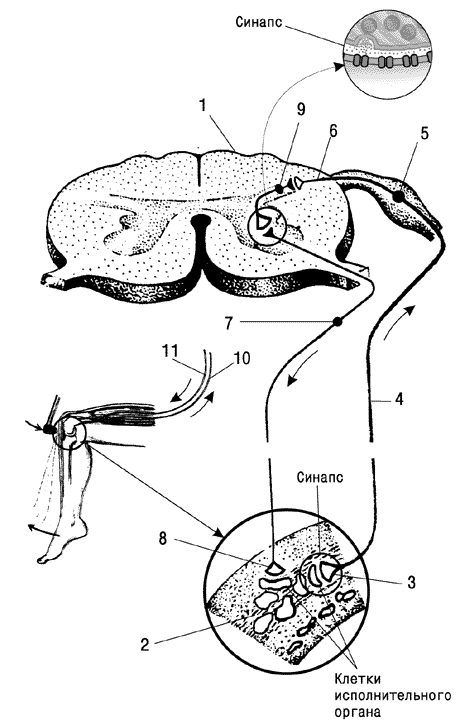
Рис. 5. Схема рефлекторной дуги.
1 – спинной мозг (поперечный срез); 2 – ткань (мышца и т.п.); 3 – окончание чувствительного нейрона; 4 – отросток чувствительного нейрона; 5 – тело чувствительного нейрона; 6 – отросток чувствительного нейрона; 7 – двигательный нейрон; 8 – окончание двигательного нейрона в органе; 9 – вставочный нейрон; 10 – чувствительное нервное волокно; 11 – двигательное нервное волокно.
Одним из фундаментальных свойств ЦНС является способность к торможению. Явление центрального торможения было открыто в 1863 г. И. М. Сеченовым, обнаружившим возникновение торможения спинальных центров лягушки при раздражении структур таламуса кристаллами соли. Впоследствии торможение изучали Н. Е. Введенский, Ч. Шеррингтон, братья Вебер, Р.М. Экклс и др. Важным шагом в выяснении природы центрального торможения оказалось выявление самостоятельного значения торможения для работы нервных центров.
Торможение – это самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. Как и процесс возбуждения, торможение является активным процессом и, соответственно, ответной реакцией. В отличие от процесса возбуждения, который может проявляться в двух основных формах – распространяющихся потенциалов действия и локальных потенциалов, торможение развивается только в форме локального процесса, и связано с существованием специфических вставочных тормозных нейронов (клетки Дейла и Реншоу). В онто- и филогенетическом аспекте торможение является более молодым свойством нервной системы, чем возбуждение.
Центральное торможение выполняет две главные функции: охранительную и координационную.
В зависимости от расположения тормозных синапсов, вида медиатора и хеморецепторов различают следующие виды торможения в ЦНС: пресинаптическое и постсинаптическое.
Пресинаптическое торможение. Одна из коллатералей, чаще всего, афферентного нейрона подходит к вставочному тормозному нейрону (клетке Дейла и др.). Аксон последнего образует синапс на другой терминали возбуждающего нейрона. В таком аксональном синапсе возникает деполяризация, которая блокирует прохождение импульсов по этой ветви афферентного нейрона. Предполагается, что деполяризация вызывает уменьшение амплитуды ПД, приходящего в возбуждающее окончание, а это уменьшает количество выделяемого им медиатора, вследствие чего амплитуда возбуждающего постсинаптического потенциала падает. Пресинаптическое торможение особенно эффективно при обработке многочисленной информации, поступающей к нейрону. В этом случае возбуждение, поступающее по одному из синаптических выходов, может быть избирательно уменьшено или полностью подавлено. Это способствует увеличению гибкости ответной реакции нейронов. Пресинаптический вид торможения обнаружен в различных отделах ЦНС, но наиболее часто он встречается в структурах мозгового ствола и спинного мозга.
Постсинаптическое торможение. При этом виде торможения тормозной вставочный нейрон контактирует с иннервируемым нейроном и вызывает гиперполяризацию на постсинаптической мембране. В этом случае резко понижается возбудимость нейрона и он не отвечает (или отвечает слабо) на любые сигналы, поступающие с других синапсов. Создаются условия для уменьшения дивергенции. Этот тип тормозных нейронов (клетки Реншоу и др.) был зарегистрирован в промежуточных нейронах спинного и продолговатого мозга, ретикулярной формации, коры больших полушарий, мозжечка и таламических ядрах.
Торможение в нервных цепях может носить характер реципрокного, прямого и возвратного. Реципрокное торможение возникает без предшествующего возбуждения, сразу. Коллатерали одного и того же афферентного нейрона контактируют с возбуждающими и тормозящими вставочными нейронами. Объединение мотонейронов на синергисты и антагонисты лежит в основе координационного акта (ходьба, бег, чередование вдоха и выдоха и т.д.). Реципрокное торможение облегчает осуществление рефлекторных актов. Возвратное торможение возникает чаще в мотонейроне по принципу обратной отрицательной связи.
Центральная нервная система координирует деятельность всех органов и систем, обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует целенаправленное поведение. Эти жизненно важные задачи решаются благодаря интегративной деятельности ЦНС.
Интегративная деятельность ЦНС –это ее способность объединять, обобщать все поступающие сигналы, отрабатывать их в связи с прошлым опытом. В результате формируется определенная реакция организма биологически или социально наиболее важная в данной ситуации.
В интегративной деятельности условно выделяют 4 основных уровня:
1. Интеграция на уровне рецептора. Рецептор, воспринимая информацию, осуществляет ее первичный отбор по интенсивности, продолжительности, модальности и формирует нервные импульсы.
2. Интегративная деятельность нейрона – это способность нейрона воспринимать возбуждение и торможение, обрабатывать их с учетом генетической и приобретенной памяти нейрона и вырабатывать временную последовательность потенциалов действия. Интегративная деятельность нейрона базируется на конвергентных свойствах нейрона и его структурных изменениях, лежащих в основе обучения и памяти.
3. Интеграция на уровне центра. Нервный центр – это совокупность нервных клеток, расположенных на различных уровнях ЦНС и обеспечивающих определенную физиологическую реакцию организма. Иерархический принцип строения нервных центров создает возможность тонко дифференцировать ответные реакции.
4. Межцентральная интеграция обеспечивает согласование деятельности различных нервных центров и формирование сложных поведенческих, эмоциональных и адаптивных реакций, организовывая деятельность организма как единого целого.
В естественных условиях любой рефлекторный акт является результатом интегративной деятельности. В основе интегративной деятельности ЦНС лежат механизмы координации.
Координация – это согласованное взаимодействие процессов возбуждения и торможения в ЦНС. Координация процессов в нервных центрах происходит при осуществлении любого рефлекторного акта. Этот процесс базируется на принципах конвергенции, дивергенции и обратной связи.
Конвергенция – схождение различных путей (тормозящих, возбуждающих) проведения нервных импульсов на одной нервной клетке. Это обуславливает интегративную функцию нейрона. Принцип конвергенции лежит в основе таких процессов, как общий конечный путь, пространственная суммация и окклюзия (см. ниже).
Дивергенция – это способность нейрона устанавливать многочисленные синаптические связи с другими нервными клетками. Благодаря процессу дивергенции один нейрон может участвовать в различных нервных реакциях и контролировать большое число других нейронов, а также каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Обратные связи – поступление нервных импульсов в ЦНС с иннервируемого органа или клетки. Обратные связи разделяют на центральные (кольцевой тип связи между нейронами) и рефлекторные (импульсы возвращаются в нервный центр с рецепторов иннервируемого органа). По эффекту обратные связи могут быть положительными и отрицательными.
В нервные центры от рецепторов обычно поступает ритмическая импульсация. При этом ответная реакция ЦНС не всегда линейно зависит от силы и частоты раздражителя. В нервных центрах можно наблюдать явление суммации допороговых стимулов и окклюзии сверхпороговых.
Суммация. Различают пространственную и последовательную суммацию. Последовательная суммация возникает при ритмической стимуляции одного рецептивного поля. В основе ее лежит механизм облегчения. Пространственная суммация допороговых стимулов происходит при одновременной стимуляции различных рецептивных полей. Она базируется на принципах облегчения и конвергенции.
Окклюзия- это процесс, при котором общая ответная реакция нервных центров на сверхпороговые стимулы меньше, чем алгебраическая сумма раздельных эффекторов каждого. Последовательная окклюзия происходит при быстрой повторной стимуляции нейрона. При этом нейрон не воспроизводит все поступающие к нему сигналы, в результате чего происходит уменьшение сильных сигналов.
В основе пространственной окклюзии лежит процесс конвергенции, который приводит к уменьшению количества суммарнореагирующих нейронов.
Различные рефлекторные реакции могут взаимодействовать между собой. Примером такого взаимодействия являются феномены иррадиации возбуждения, доминанта и принцип общего конечного пути.
Общий конечный путь. Данный принцип введен в физиологию Ч. Шеррингтоном и основан на способности различных проводящих путей создавать синаптические контакты на одной и той же эффекторной клетке. В основе этого лежит принцип конвергенции. К мотонейронам спинного мозга кроме первичных афферентных волокон конвергируют волокна различных нисходящих трактов, идущих из центральных структур мозга, а также аксоны возбуждающих и тормозных вставочных нейронов спинного мозга. Вследствие этого Ч. Шеррингтон именно мотонейроны рассматривал как общий конечный путь многочисленных структур мозга, связанных с регуляцией моторных функций. Принцип общего конечного пути показывает, каким образом одна и та же конечная реакция может быть получена при раздражении различных структур мозга. Этот принцип имеет важное значение для анализа рефлекторной деятельности нервной системы.
Иррадиация возбуждения – это широкое распространение возбуждения по различным нервным центрам. В основе этого процесса лежит большая продолжительность и сила поступающих стимулов в ЦНС, высокая возбудимость нейронов и ослабление центрального торможения. Механизм иррадиации – дивергенция.
Иррадиация возбуждения по нервным центрам способствует возможности одних нейронов участвовать в различных нервных реакциях и контролировать деятельность других нейронов. Однако иррадиация возбуждения может стать патологической в связи с возникновением сильного очага возбуждения и с изменением свойств нервной ткани, усиливающим распространение возбуждения по ней, как это бывает при эпилепсии.
Доминанта – временно господствующий очаг возбуждения в ЦНС, обуславливающий интегральный характер функционирования нервных центров в каждый данный период времени и определяющий целесообразное поведение человека.
Доминантный очаг возбуждения притягивает к себе возбуждение из других нервных центров и одновременно подавляет их деятельность. Доминантный очаг обладает также и свойством притягивания сигналов с других рецептивных полей.
Доминанта может исчезнуть при возникновении более сильной доминанты, реализации доминантного рефлекса или ее затухания в следствие истощения энергетических ресурсов.
Различают следующие виды доминанты: физиологическую и патологическую. Физиологическая доминанта обусловлена биологическими и социальными потребностями (мотивами). Патологическая – проявляется в виде вредных привычек (табакокурение, алкоголизм, наркомания) или при психических расстройствах.
И. П. Павлов указывал также на то, что принцип доминанты лежит в основе формирования временной функциональной связи (условного рефлекса). Таким образом, явление доминанты является одним из важнейших принципов работы ЦНС.
Тема 3. Частная физиология центральной нервной системы.
Вопросы для самоподготовки.
I. Спинной мозг.
1. Строение спинного мозга (расположение белого и серого вещества, понятие «сегмент», отделы, оболочки).
2. Функции спинного мозга.
2.1. Проводниковая функция (проводящие пути спинного мозга).
2.2. Рефлекторная функция. Рефлексы спинного мозга.
2.3. Спинальный шок.
II. Продолговатый мозг.
1. Строение продолговатого мозга (границы, пирамиды, оливы, особенности расположения белого и серого вещества, ядра продолговатого мозга).
2. Функции продолговатого мозга.
2.1. Проводниковая функция продолговатого мозга.
2.2. Рефлекторная деятельность продолговатого мозга:
- центры жизненноважных рефлексов (дыхательный, сердечно-сосудистый);
- защитные рефлексы (мигание, чихание, кашель, рвотный акт и др.);
- рефлексы, связанные с пищеварительной деятельностью (глотание, отделение пищеварительных соков и др.);
- рефлексы, связанные с функциями ядер черепно-мозговых нервов, расположенных в продолговатом мозге (в том числе и вегетативные);
3. Участие продолговатого мозга в регуляции мышечного тонуса и рефлексов позы.
4. Ретикулярная формация ствола мозга:
- строение РФ (расположение, ядра РФ, афферентные и эфферентные связи, виды нейрональных контактов);
- функции РФ (контроль сна и бодрствования, участие в регуляции вегетативных функций, фазный и тонический мышечный контроль, участие в механизмах формирования целостных условно -рефлекторных реакций организма).
III. Средний мозг.
1. Строение среднего мозга (границы, ядра).
2. Функции среднего мозга.
2.1. Проводниковая функция.
2.2.Рефлекторная деятельность среднего мозга:
- роль ядер бугров четверохолмия («старт-рефлексы»);
- роль красных ядер и черной субстанции в распределении мышечного тонуса (синдром Паркинсона и децеребрационная ригидность).
3. Роль среднего мозга в сохранении нормального положения тела в пространстве (выпрямительные и статокинетические рефлексы).
IV. Мозжечок
1. Строение мозжечка (червь, полушария, кора и белое вещество, ножки, ядра; связи мозжечка с другими структурами ЦНС).
2. Функции мозжечка:
- участие в координации движений;
- регуляция мышечного тонуса;
- сохранение позы и равновесия тела;
- участие в регуляции вегетативных функций (функций внутренних органов);
- влияние мозжечка на образование условных рефлексов.
3. Симптомы нарушения функций мозжечка (астазия, атаксия, астения, атония, дистония и др.).
V. Промежуточный мозг.
1. Составные части промежуточного мозга (эпиталамус, метаталамус, таламус и гипоталамус).
2. Таламус.
2.1. Нейронная организация.
2.1. Функции таламуса:
- роль специфических (переключательных и ассоциативных) ядер, моторных и неспецифических ядер;
- регуляция важных физиологических состояний (смена сна и бодрствования, сохранение сознания, развитие процессов внутреннего торможения и др.).
3. Гипоталамус.
3.1.Нейронная организация.
3.2. Афферентные и эфферентные связи гипоталамуса.
3.3. Функции гипоталамуса:
- роль гипоталамуса в регуляции вегетативных функций;
- участие в регуляции поведенческих реакций;
- гипоталамо-гипофизарная система.
3.4. Функциональные расстройства у людей с повреждениями гипоталамуса.
VI. Базальные ядра.
1. Структуры, входящие в состав базальных ядер и их связи.
2. Функции базальных ядер:
- обеспечение миостатических реакций (плавности движений);
- обеспечение автоматизма движений – бессознательного их выполнения;
- обеспечение движения мимических мышц и участие в формировании эмоциональных реакций;
- формирование защитных ориентировочных рефлексов.
3. Патофизиология базальных ганглиев:
- гипофункция медиаторных систем (болезнь Паркинсона);
- гиперфункциональные симптомы (ригидность, баллизм, атетоз, хорея, тремор).
VII. Кора больших полушарий.
1. Организация коры больших полушарий (клеточные слои, доли, области, поля).
2. Древняя и старая кора.
2.1. Структуры, входящие в состав (обонятельный мозг и лимбическая область).
2.2. Функции древней и старой коры:
- обеспечение реакций настораживания и внимания;
- регуляция вегетативных функций;
- осуществление видоспецифического (инстинктивного) поведения;
- формирование эмоций;
- осуществление социального поведения;
- участие в процессах сохранения памяти.
3. Функции новой коры.
3.1.Чувствительные зоны коры большого полушария.
3.2.Моторные зоны коры большого полушария.
3.3.Электрические явления в коре больших полушарий (электроэнцефалография – ЭЭГ).
Основополагающий материал.
Спинной мозг представляет собой низший и филогенетически наиболее древний отдел ЦНС. Спинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. От каждого спинномозгового сегмента отходят две пары передних и задних корешков. Задние корешки образованы центральными отростками афферентных нейронов, тела которых находятся в спинномозговых ганглиях. В передних корешках проходят аксоны α – и γ– мотонейронов, а также преганглионарных нейронов ВНС. Такое распределение афферентных и эфферентных волокон получило название закона Белла – Мажанди. После перерезки передних корешков происходит нарушение двигательной функции определенных мышц. При перерезке же задних корешков происходит потеря чувствительности определенного участка тела. Двигательная функция при этом не нарушается.
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются как соматические, так и вегетативные функции организма. К числу наиболее простых рефлекторных реакций относятся сухожильные рефлексы и рефлексы растяжения. Более сложно организованы сгибательные и разгибательные рефлексы, ритмические и позные рефлексы, к которым относятся чесательный, потирательный и шейные тонические рефлексы. Важной функцией мотонейронов спинного мозга является поддержание тонуса мышц (постоянное слабое их напряжение и незначительное укорочение). Благодаря тонусу все рефлекторные двигательные акты протекают мягко и плавно. Но тонус центров спинного мозга регулируется высшими отделами ЦНС. В крестцовом же отделе спинного мозга располагаются центры актов дефекации, мочеиспускания и половой деятельности.
Спинной мозг обеспечивает также проводниковую функцию. Белое вещество, образованное мякотными волокнами, образует систему проводящих путей, где каждый тракт занимает вполне определенное положение. Более подробно о проводящих путях изложено в методическом пособии «Анатомия ЦНС».
При поражении спинного мозга наблюдается возникновение спинального шока, который характеризуется уменьшением или исчезновение рефлекторной активности ниже места повреждения. Длительность и сила его зависят от уровня филогенетического развития животного и от уровня повреждения спинного мозга (чем выше уровень повреждения, тем тяжелее протекает спинальный шок). Существует два основных предположения относительно развития спинального шока. Согласно одному шок возникает вследствие выпадения преимущественно возбуждающего влияния со стороны высших центров на активность различных спинальных уровней ниже места повреждения. Согласно второму – перерезка устраняет подавляющее влияние высших моторных центров на спинальное торможение.
Продолговатый мозг. Продолговатый мозг посредством черепно–мозговых нервов связан с разнообразными рецепторами и эффекторами организма, а через восходящие и нисходящие проводящие пути – с другими отделами ЦНС.
Продолговатый мозг осуществляет рефлекторную и проводниковую функции. Рефлексы продолговатого мозга сложнее и разнообразнее рефлексов спинного. Функции продолговатого мозга имеют жизненно важное значение: в нем находятся дыхательный и сосудистый центры; продолговатый мозг оказывает вегетативное рефлекторное влияние на внутренние органы и сосуды (Х пара ч.м.н.). Кроме того, продолговатый мозг осуществляет ряд защитных рефлексов (мигание, чихание, кашель, рвотный акт и др.), ряд рефлексов, связанных с пищеварительной деятельностью (глотание, отделение пищеварительных соков и др.), и тонические рефлексы на скелетную мускулатуру.
Большой интерес вызывают рефлексы позы, осуществляемые с уровня продолговатого мозга.
В процессе эволюции у животного и человека выработалась нормальная поза, характеризующаяся положением головы теменем вверх. Поэтому у организма возникла и закрепилась группа тонических рефлексов, направленных на сохранение равновесия при угрозе его нарушения и на восстановление нормальной позы в тех случаях, когда она уже нарушена. Рефлексы позы возникают при изменении положения головы в пространстве и по отношению к туловищу. Так, при наклоне или при повороте головы в ту или другую сторону можно наблюдать рефлекторное увеличение тонуса разгибателей с соответствующей стороны и снижение его на противоположной. При опускании головы животного происходит снижение разгибательного тонуса передних конечностей при некотором повышении тонуса задних. При подъеме головы кверху происходит повышение тонуса мышц-разгибателей передних конечностей и снижение – на задних.
Рефлексы позы осуществляются с рецепторов вестибулярного аппарата, с проприорецепторов мышц и сухожилий шеи, а также с экстерорецепторов кожи шеи.
Отчетливое проявление позных рефлексов можно наблюдать на больных людях при нарушении у них деятельности вышележащих отделов мозга. У здорового человека при изменении положения головы методом рентгенокимографии было обнаружено повышение тонуса диафрагмы с той стороны, куда была повернута голова.
Средний мозг. Средний мозг состоит из четверохолмия и ножек мозга. Связи среднего мозга с другими отделами нервной системы очень богаты. Через средний мозг проходят различные восходящие пути к таламусу и мозжечку, нисходящие пути из коры больших полушарий, полосатого тела, гипоталамуса к самому среднему мозгу и к ядрам продолговатого и спинного мозга.
В среднем мозге замыкается ряд рефлексов. Так, при участии нейронов бугров четверохолмия осуществляются ориентировочные зрительные и слуховые рефлексы («старт–рефлексы»).
Черная субстанция участвует в сложной координации движений. Ее нейроны, содержащие дофамин, своими аксонами связаны с передним мозгом (участие в регуляции эмоционального поведения), и ядрами полосатого тела (контроль сложных двигательных актов). Повреждения черной субстанции приводят к нарушению тонких движений пальцев рук, развитию мышечной ригидности и тремору (болезнь Паркинсона).
В составе среднего мозга также имеются важные эффекторные образования – красные ядра. Они принимают значительное участие в осуществлении мышечного тонуса и нормальной позы тела. При перерезке ствола мозга ниже уровня красного ядра у животных наблюдается резкое повышение тонуса мышц разгибателей. Конечности при этом сильно вытянуты, голова запрокинута назад, хвост приподнят. Это состояние называется децеребрационная ригидность. Развитие децеребрационной ригидности связано с выключением импульсации, поступающей в спинной мозг по кортико–, ретикуло– и руброспинальному трактам, которые активируют мотонейроны мышц–сгибателей. При этом начинает преобладать активность вестибулоспинальной системы, повышающей тонус мотонейронов мышц–разгибателей. Децеребрационная ригидность наблюдается и у человека при травмах среднего мозга.
Средний мозг осуществляет также ряд установочных тонических рефлексов – выпрямительных и статокинетических (см. таблицу 2).
Таблица 2.

Ретикулярная формация – протяженная структура в стволе мозга – важная интегративная область неспецифической системы. Это образование связано со многими отделами ЦНС. В РФ идут афференты в составе спиноретикулярного тракта, пути от всех черепно–мозговых нервов, и практически от всех чувствительных путей. Дополнительная афферентация поступает от многих других отделов головного мозга – от моторных и сенсорных областей коры, от таламуса и гипоталамуса. Во множестве имеются также эфферентные связи – нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре, гипоталамусу и лимбической системе.
Функции РФ еще не вполне изучены. Считается, что она участвует в ряде процессов:
– регуляция возбудимости коры: уровня осознания стимулов и реакций, ритма сон–бодрствование (восходящая активирующая ретикулярная система);
– придание аффективно–эмоциональной окраски сенсорным стимулам, особенно болевым, за счет передачи афферентной информации в лимбическую систему;
– двигательная регуляция функций, в том числе жизненно важных рефлексов (кровообращения, дыхания, глотания, кашля и чихания), при которых должны взаимно координироваться разные афферентные и эфферентные системы;
– участие в регуляции позных и целенаправленных движений в качестве важного компонента двигательных центров ствола мозга.
Мозжечок. Строение мозжечка подробно изложено в нашем предыдущем методическом пособии («Анатомия ЦНС»), поэтому здесь мы не будем останавливаться на нем.
Многое из того, что известно о функциях мозжечка, связано с наблюдениями неврологов–клиницистов. Больные с его повреждениями не парализованы и у них не нарушена чувствительность, хотя мозжечок получает в основном сенсорные сигналы и дает проекции к двигательным центрам. Симптоматика мозжечковых патологий характеризуется аномальным выполнением движений и исчезновением сопряжения между ними и позой. Следовательно, мозжечок участвует в сенсомоторной интеграции и выполняет важную функцию координации движения.
Функциональная двойственность мозжечка соответствует его структурной и филогенетической двойственности. То есть, его филогенетически более древние медиальные структуры (червь) получают спинальные, вестибулярные и зрительные сигналы и тесно связаны с двигательной системой ствола и спинного мозга. К развившимся позже латеральным структурам (полушариям) информация идет прежде всего от коры мозга, к которой от них в свою очередь направляются восходящие пути. Эти особенности приводят к выводу, что червь осуществляет главным образом регуляцию и координацию движений в ходе их выполнения, а его полушария больше участвуют в подготовке, программировании движений. Кроме того, недавно выявлены долговременные изменения синаптической передачи сигналов в мозжечке. Полагают, что такая пластичность играет роль в долговременной двигательной адаптации и двигательном научении.
Связь мозжечка с ретикулярной формацией ствола мозга, по–видимому, обеспечивает его висцеральные (адаптационно–трофические) функции, а с корой больших полушарий – влияние на образование условных рефлексов.
Основные функции мозжечка определяют и характер патологических симптомов при нарушении его деятельности.
Атония – характеризуется ослаблением мышечного тонуса и неспособностью поддерживать определенную позу. Обычно атония сопровождается симптомом астении, которая характеризуется слабостью и быстрой утомляемостью мышц. Очевидно, атония и астения связаны с выпадением функции мозжечка контролировать познотонические рефлексы.
Астазия – проявляется в способности мышц давать колебательные и дрожательные движения. Мышечный тремор особенно выражен в начале и конце движения, что в значительной мере препятствует завершению целенаправленного движения.
Асинергия – страдают содружественные движения, происходит как бы распад программы движения и целостное движение состоит не из одновременных содружественных актов, а из последовательности ряда простых движений. Так, например, касание кончика носа мозжечковый больной осуществляет в три приема. Асинергия сочетается с дисметрией, или утратой соразмерности движения. Движения становятся преувеличенно размашистыми и теряют свою точность, в результате чего цель достигается только после многократных ошибок.
У мозжечковых больных деформируется походка – атаксия. Больным трудно стоять и ходить, особенно в темноте; им приходится хвататься за что–нибудь руками, походка становится шатающейся, как будто в состоянии опьянения. Для них также характерно головокружение, тошнота и рвота.
Серьезные трудности возникают при попытках быстро «изменять программу», например быстро поворачивать руку то ладонью вверх, то ладонью вниз. Такие движения совершаются неритмично и медленно (адиадохокинез). Нарушена также координация мышц лица, необходимая для четкого произнесения слов (дизартрия). Речь становится медленной, невыразительной, монотонной.
Промежуточный мозг. Топографически и функционально промежуточный мозг подразделяется на эпиталамус, таламус и гипоталамус.
Эпиталамус, или надталамическая область, состоит из расположенного под мозолистым телом свода и из железы внутренней секреции эпифиза.
Таламус, или зрительный бугор (парный орган), представляет собой скопление серого вещества. Он является, пожалуй, единственным коллектором различного вида чувствительности, идущей в кору больших полушарий, а также в другие образования ЦНС. Нервные клетки таламуса группируются в большое количество ядер (до 60), которые топографически разделяют на переднюю, медиальную и латеральную группы. По функции таламические ядра можно дифференцировать на специфические (переключательные и ассоциативные) , неспецифические и моторные.
Для специфических (проекционных) ядер характерна отчетливая топографическая и функционально разграниченная проекция к различным областям коры мозга, по которой с периферии поступают раздражения определенной модальности. Что касается ассоциативных ядер, то они не могут быть отнесены к какой–либо одной сенсорной системе и получают афферентные импульсы от специфических проекционных ядер. Три ядра этой группы имеют связи с главными ассоциативными областями коры (теменной, височной, лобной), а четвертое – с лимбической корой больших полушарий и гипоталамусом (мотивационные влияния). По–видимому, ассоциативные ядра участвуют в высших интегративных процессах. Неспецифические таламические ядра не связаны с проведением раздражения какой–либо одной модальности. К ним приходит афферентация в основном из ретикулярной формации мозгового ствола, и они обнаруживают диффузные проекции во все области коры. Этой структурной особенностью обусловлена их способность регулировать возбудимость корковых нейронов – усиливать (реакция вовлечения) или угнетать разряды корковых нейронов. Двигательное моторное ядро участвует в регуляции движений, так как имеет вход от мозжечка и базальных ганглиев, и одновременно дает проекции в моторную зону коры больших полушарий. При перерезке мозга ниже таламуса происходит нарушение локомоции и сложных двигательных рефлексов типа глотания, жевания и сосания. Кора, в свою очередь, может оказывать тормозные и облегчающие влияния на таламические ядра, т.к. между корой и таламусом имеются двусторонние циклические связи. Поэтому, не случайно, таламокортикальная система связана с регуляцией таких физиологически важных состояний, как смена сна и бодрствования, сохранение сознания, развитие процессов внутреннего торможения, в условнорефлекторной деятельности и поведении.
Таламус также является высшим подкорковым центром болевой чувствительности (неспецифические ядра). При некоторых повреждениях таламуса появляются мучительные болевые ощущения, которые могут быть спровоцированы прикосновением к коже, звуковым или световым раздражением. Иногда при поражении таламуса может наступить наоборот, состояние полной аналгезии. Таким образом, таламус, являясь коллектором практически всех чувствительных путей, принимает участие в формировании ощущений, в механизме эмоций и психорефлексов (смех, плач).
Гипоталамус– филогенетически старый отдел промежуточного мозга, который играет важную роль в поддержании постоянства внутренней среды организма и обеспечении интеграций функций вегетативной, эндокринной и соматической систем.
В гипоталамусе можно выделить несколько десятков ядер, большинство из которых имеют плохо очерченные границы и, за малым исключением, их трудно рассматривать как центры с узкой локализацией специфических функций. Более перспективным является деление гипоталамуса на области и зоны, обладающие определенной функциональной спецификой. Так, например, некоторые ядра преоптической и передней групп объединяются в гипофизотропную область, нейроны которой продуцируют рилизинг–факторы (либерины) и ингибирующие факторы (статины), которые регулируют деятельность передней доли гипофиза (аденогипофиз).
Срединная группа ядер формирует медиальный гипоталамус, где имеются нейрорецептивные клетки, реагирующие на различные изменения внутренней среды организма (температуру крови, водно-электролитный состав плазмы, содержание гормонов в крови). Посредством нервных и гуморальных механизмов медиальный гипоталамус управляет деятельностью гипофиза.
Латеральный гипоталамус представляет собой безъядерную зону, в которой сосредоточены проводниковые элементы, формирующие пути к верхним и нижним отделам стволовой части мозга. Нервные клетки в латеральном гипоталамусе расположены диффузно.
В целом гипоталамус имеет богатые двусторонние (афферентные и эфферентные) связи с таламусом и через передние таламические ядра – с корой больших полушарий; афферентный вход от лимбической системы мозга и ретикулярной формации среднего мозга; гипоталамус связан с базальными ганглиями переднего мозга и мозжечком. Эфферентные связи гипоталамуса с вегетативными и соматическими ядрами головного и спинного мозга осуществляются через полисинаптические пути РФ ствола мозга.
Благодаря своим обширным связям со многими структурами ЦНС гипоталамус обеспечивает стыковку нервного, эндокринного и гуморального механизмов регуляции деятельности организма, результатом чего является его участие в регуляции вегетативно–висцеральных функций, направленных на обеспечение различных форм деятельности (пищевое и сексуальное поведение, процессы сохранение вида) в регуляции систем, обеспечивающих сон и бодрствование, внимание, эмоциональную сферу, процессы памяти (т.е. в осуществлении сомато–вегетативной интеграции).
У человека нарушения деятельности гипоталамуса бывают связаны, главным образом, с опухолевыми, травматическими или воспалительными поражениями. У таких больных наблюдаются сложные функциональные расстройства (табл.3). При этом расстройства восприятия, памяти и цикла сон–бодрствование частично связаны с повреждением восходящих и нисходящих путей, соединяющих гипоталамус с лимбической системой.
Таблица 3.