Правило десяти процентов. Правило одного процента
В соответствии с законом пирамиды энергий, с одного трофического уровня экологической пирамиды переходит на другой ее уровень в среднем не более 10% энергии — этоправило десяти процентов. Закон пирамиды энергий позволяет делать расчеты необходимой земельной площади для обеспечения населения продовольствием и другие эколого-экономические подсчеты. Среднемаксимальный переход энергии (или вещества в энергетическом выражении) с одного трофического уровня экологической пирамиды на другой, составляя 10%, может колебаться в пределах 7—17%. Превышение этой величины недопустимо, иначе могут произойти полные исчезновения популяций.
Правило одного процента — изменение энергетики природной системы в пределах одного процента выводит природную систему из равновесного (квазистационарного) состояния. Эмпирически это правило подтверждается исследованиями климата и других природных процессов.
Все крупномасштабные явления на поверхности Земли (мощные циклоны, извержения вулканов, процесс глобального фотосинтеза), как правило, имеют суммарную энергию, не превышающую 1% от энергии солнечного излучения, падающего на поверхность нашей планеты. Переход энергетики процесса за это значение обычно приводит к существенным аномалиям — резким климатическим отклонениям, переменам в характере растительности, крупным лесным и степным пожарам.
Как и в случае правила десяти процентов, многое зависит от состояния природной системы, в которой происходят изменения. Это делает данное правило вероятным, дает лишь ориентиры, которым целесообразно следовать или учитывать возможную с большой вероятностью цепь событий, связанных с выходом системы из равновесного (квазистационарного) состояния.
Особое значение правило данного процента имеет для глобальных систем. Их энергетика, как предполагают, принципиально не может превзойти уровень примерно 0,2% от поступающей солнечной радиации (уровень энергетики фотосинтеза) без катастрофических последствий. Вероятно, это непреодолимый и недопустимый для человечества порог (из него следует и "ядерная зима").
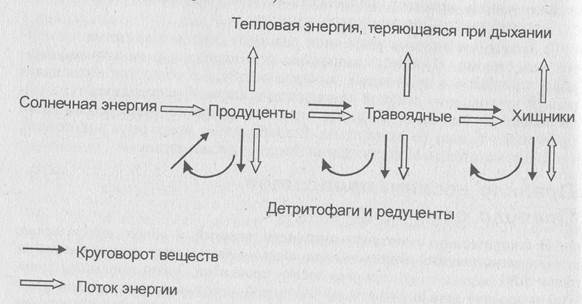
Рис. 2. Тепловая энергия, теряющаяся при дыхании
Трофические цепи, которые начинаются с фотосинтезирующих организмов, называютцепями выедания (пастбищными, цепями потребления).
Цепи, которые начинаются с отмерших остатков растений, трупов и экскрементов животных— детритные цепи разложения.
Трофические цепи не изолированы друг от друга; тесно переплетаясь, они образуют трофические сети. Благодаря трофическим связям в экосистеме происходит трансформация биогенных веществ и аккумуляция энергии с последующим распределением их между видами и популяциями. Чем богаче видовой состав, тем разнообразнее направление и скорость потоков энергии в экосистеме.
Трофические цепи питания основываются на:
- втором законе термодинамики, согласно которому некоторая часть энергии рассеивается и становится недоступной для использования в виде тепловой энергии;
- а также на закономерностях поедания организмов, которые заключаются в том, что по мере продвижения по трофической цепи хищников животные увеличиваются в размерах, но число их уменьшается. И наоборот, в трофических цепях паразитов организмы уменьшаются в размерах, а их численность увеличивается.
В экосистемах разных типов мощность потоков энергии через цепи выедания и разложения различна:
- в водных сообществах часть энергии, фиксированной одноклеточными водорослями, поступает к питающимся фитопланктоном животным, далее к хищникам и уже меньшая часть включается в цепи разложения;
- в большей части экосистем суши наблюдается обратное соотношение. Так, в лесах более 90% ежегодного прироста растительной массы поступает через опад в детритные цепи.
Число звеньев в цепи питания может быть различным, но в основном их обычно бывает от 3 до 5.
Совокупность организмов, объединенных определенным типом питания, носит название"трофический уровень". Различают:
- первый уровень, который занимают автотрофы (продуценты);
- второй — растительноядные животные (консументы первого порядка);
- третий - хищники, питающиеся травоядными животными (консументы второго порядка), и паразиты первичных консументов;
- четвертый трофический уровень образуют вторичные хищники (консументы третьего порядка) и паразиты вторичных консументов.
Трофических уровней может быть и больше, когда учитываются паразиты, живущие на консументах предыдущих уровней.
Примером цепи питания может служить цепь питания биологического биоценоза.
Например, начинается цепь с улавливания солнечной энергии : цветком. Бабочка, питающаяся нектаром цветка, представляет собой второе звено в этой цепи. Стрекоза нападает на бабочку. А спрятавшаяся лягушка ловит стрекозу, но сама является добычей для ужа, который в свою очередь будет съеден ястребом. Цепь питания замкнулась. Потенциальным (но не обязательным) заключительным звеном пищевой цепи является человек.
Все рассмотренные выше процессы связаны с синтезом и трансформацией органического вещества в трофических сетях и характеризуют "пастбищные цепи".
"Детритные цепи" начинаются с разложения мертвой органики особыми группами консументов — сапрофагами. Сапрофаги механически, а отчасти и химически, разрушают мертвое органическое вещество, подготавливая его к воздействию редуцентов. В наземных экосистемах этот процесс (по большей части) проходит в подстилке и в почве. Активное участие в разложении мертвого органического вещества принимают почвенные беспозвоночные животные (членистоногие, черви) и микроорганизмы. Процесс деструкции идет последовательно, сапрофаги меняют друг друга в соответствии со спецификой видового питания. Механическое разрушение производят крупные сапрофаги (например, насекомые), а процесс минерализации осуществляют уже другие организмы (прежде всего бактерии и грибы).
Поскольку сообщества сапрофагов отличаются относительно слабой жесткостью организации, то в детритных цепях идут стохастические процессы формирования сапрофагов, отдельные их виды легко заменяются другими видами, велика роль внешних факторов среды обитания и конкурентного исключения (Н.М. Чернова, Н.А. Кузнецова, 1986). То есть с уровня консументов поток органического вещества идет по разным группам потребителей: живое органическое вещество идет по "пастбищным цепям", а мертвое — по "детритным цепям".
Продуктивность экосистемы
Зависимость между продуктивностью и климатическими характеристиками. Все организмы нуждаются для построения своих тел в веществе, а для поддержания своей жизнедеятельности — в энергии. Солнечный свет, двуокись углерода, вода и минеральные соли — это ресурсы, необходимые для создания первичной продукции. На скорость фотосинтеза оказывает существенное влияние и температура. Качество и количество света, наличие воды и биогенных элементов, а также температура — весьма изменчивые факторы, которые способны лимитировать первичную продукцию.
На каждый квадратный метр земной поверхности ежеминутно попадает от 0 до 5 Дж солнечной энергии. По спектральному составу только около 44% падающего коротковолнового света пригодно для синтеза, а значительная доля солнечной энергии растениям недоступна. Наиболее высокой эффективностью использования солнечной энергии обладают хвойные леса: 1-3% физиологически активной 'радиации (ФАР) они превращают в биомассу. Листопадные леса превращают в биомассу только 0,5-1% ФАР, а пустыни еще меньше - 0,01-0,02%. Максимальная эффективность фотосинтеза зерновых культур при идеальных условиях не превышает 3-10%.
Использование доступного для растения света немного улучшается при хорошей обеспеченности и другими ресурсами.
Вода - незаменимый ресурс и как составная часть клетки, и как участник фотосинтеза. Потому продуктивность всегда тесно связана с количеством выпадающих осадков.
На продуктивность существенное влияние оказывает и температура среды. Эта зависимость носит сложный характер.
Продукция наземного сообщества зависит и от содержания в почве необходимых для растений различных микроэлементов. Особенно большое влияние оказывают соединения азота. Причем их происхождение должно быть биологическим, т. е. результатом фиксации азота микроорганизмами, а не геологическим.
На продуктивность существенное влияние оказывает и деятельность человека. По мере развития сельского хозяйства в направлении получения максимума продукции воздействие на природу, обусловленное перераспределением энергии и веществ на поверхности Земли, постоянно возрастает. Совершенствование орудий труда, внедрение высокоурожайных культур и сортов, требующих большого количества питательных веществ, стали резко нарушать природные процессы.
Разрушительно действуют необоснованные земледельческие приемы и системы земледелия, которые вызывают:
- эрозию почв и утрату плодородного слоя;
- засоление и заболачивание орошаемых массивов;
- снижение биологического разнообразия естественных ландшафтов;
- загрязнение поверхностных и подземных вод остатками пестицидов и нитратов;
- исчезновение диких животных в результате разрушения мест их обитания и многое другое.
Для регулирования и решения этих проблем предлагают научно обоснованные приемы и способы, позволяющие в определенных случаях лишь частично предотвратить или снизить нежелательные эффекты, возникающие при получении первичной биологической продукции. В последние десятилетия все активнее вводятся экологические ограничения.
Существует объективный природный предел - порог снижения естественного плодородия, при приближении к которому вся техническая мощь человека становится менее эффективной. Во второй половине XX в. произошло существенное увеличение первичной биологической продукции за счет внедрения новых высокоурожайных сортов зерновых культур, применения большого количества минеральных удобрений и использования средств защиты растений. Однако этот показатель перестал расти, что явилось отражением действия закона снижения энергетической эффективности природопользования.
Но численность человечества продолжает расти, а плодородной земли больше не становится. Поэтому увеличение КПД зеленых растений является наиболее насущной проблемой при решении первейших задач жизнеобеспечения человека. В табл. 4 проведен один из вариантов расчета первичной продукции земного шара по итогам исследований П. Дювиньо.
| Экосистема | Поверхность, млн км2 | Выход фотосинтеза, % | Продуктивность, т/га | Общая продуктивность органического вещества млрд т/га |
| Леса | 40,7 | 0,38 | 20,4 | |
| Степи | 25,7 | 0,1 | 1,5 | 3,8 |
| Пашни | 14,0 | 0,25 | 5,6 | |
| Пустыни | 54,9 | 0,01 | 0,2 | 1,1 |
| Антарктида | 12,7 | |||
| Океан | 0,05 | 0,8 | ||
| Всего | 60,9 |
Из данных табл. 4 видно, что экосистема океана дает половину всей продукции планеты, леса — третью часть, а пашни (вместе со степями) - около одной десятой.
При подсчете вторичной продукции экосистем производят вычисления отдельно для каждого трофического уровня, потому что при движении энергии от одного трофического уровня к другому она прирастает за счет поступления с предыдущего уровня. При изучении общей продуктивности экосистемы следует помнить, что прирост вторичной продукции всегда происходит не параллельно росту первичной, а за счет уничтожения какой-то ее части. То есть происходит как бы изъятие, вычитание вторичной продукции из общего количества первичной. Поэтому оценку продуктивности экосистем всегда проводят по первичной продукции. В целом вторичная продуктивность колеблется в пределах от 1 до 10%, а это в свою очередь зависит от свойств животных и особенностей поедаемого или корма.