Динамика численности млекопитающих
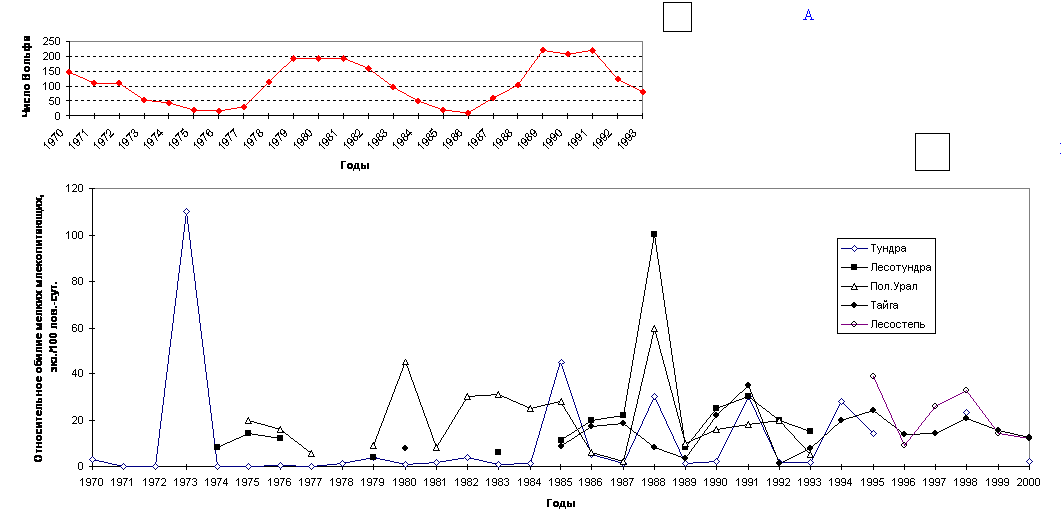
Изучение динамики численности животных само по себе, уже по определению, является мониторингом. Однако, принимая как аксиому тот факт, что динамика численности наряду с популяционными механизмами саморегуляции зависит от условий окружающей среды, мы можем рассматривать особенности ее в качестве критерия оценки этой среды, подверженной трансформациям под действием естественных или антропогенных факторов. В этом смысле динамика численности животных (годичные флуктуации (Одум, 1986)) является важным объектом экологического мониторинга не только в прикладном (для охотоведения, сельского хозяйства и т.д.), но и в теоретическом плане. В настоящей работе мы на основе собственных многолетних данных и анализе литературного (Балахонов и др., 1997 и др.) и ведомственного (Фонд..., 1992; Обзор..., 1995, 1996, 1997, 1998, 1999; Ресурсы..., 1996) материала по динамике численности млекопитающих Тюменской области постараемся осветить некоторые особенности использования этого показателя в системе экологического мониторинга (Гашев, 2000 г.). Прежде всего, нужно иметь в виду, что годовые и многолетние колебания численности млекопитающих во многом определяются естественными причинами. Так, например, В.С.Дьячков (1989), С.А.Корытин (1994) и др. опосредованно связывают их с 11-летними циклами солнечной активности (Рис. 3 а). Поэтому далеко не всегда снижение или, наоборот, рост численности того или иного вида может свидетельствовать, скажем, о каком-то действии антропогенных факторов. В этом плане хорошо известен пример изменения численности зайца-беляка и связанные с ними флуктуации рыси в Канаде во второй половине 18 - первой половине 20 века (MacLulich, 1937). Колебания численности зайца-беляка имели достаточно четкую 10-летнюю периодичность (в среднем - 9.6 года), и этим колебаниям соответствовали колебания численности рыси, питающейся зайцем-беляком. При этом пики численности зайца на год или чуть более предшествовали пикам численности рыси. Такие же регулярные естественные колебания численности характерны и для других животных. При этом многолетние флуктуации численности отдельных видов условно делят на цикличные и не цикличные, используя индекс цикличности популяций - s (Уильямсон, 1975 цит. по Кривошеев, Кривошеева, 1998). Мы предлагаем использовать интегральный материал по всей группе мелких млекопитающих. Так, численность мелких млекопитающих значительно колеблется по годам: в Тюменской области эти флуктуации прослежены более чем за 30 лет: с 1970 по 2002 гг. (Рис.14 б)(Балахонов и др., 1997, наши данные и др.). Анализируя картину в целом, можно отметить, что за этот период отмечалось 10 депрессий (1971-72; 1974; 1977-78; 1981; 1984; 1986-87; 1989; 1992-93; 1996; 1999-2000) и 10 пиков численности (1970; 1973; 1975-76; 1979-80; 1982-83; 1985; 1988; 1991; 1994-95; 1998) мелких млекопитающих. Периодичность колебаний составила около 3 лет (в среднем 2.9 года). Хорошо видно, что, несмотря на некоторые смещения сроков пиков, и спадов в разных природных зонах, общая картина сохраняется везде. Бросается в глаза, что в наиболее экстремальных условиях среды (тундра и лесотундра) выраженность колебаний выше по амплитуде. Причем, говоря о более экстремальных условиях среды в тундре и лесотундре, мы имеем в виду объективные характеристики этих условий, соглашаясь с тем, что виды, населяющие эти природные подзоны, безусловно, имеют целый ряд адаптивных приспособлений для переживания этих экстремальных условий. Сама же «экстремальность» отражена в значениях коэффициентов «вязкости» (К) и «упругости» (G) среды в нашей модели устойчивости. Образно говоря, в этих природных зонах под одним и тем же «шаром» (сообществом) плоскость прогибается незначительно, а сама поверхность «чаши» практически лишена шероховатости. В этих условиях маятникообразные колебания в системе, безусловно, больше по амплитуде. Это и наблюдается в отношении колебания численности мелких млекопитающих. При анализе этого материала мы отмечаем, что еще более правильную периодичность имеют не пики или депрессии численности мелких млекопитающих, а точки перехода кривой динамики численности через среднюю многолетнюю - это происходит строго через 2 года на третий. Выявленная закономерность, безусловно, может быть использована при прогнозировании вспышек численности мелких млекопитающих при проведении ежегодных контрольных учетов в рамках экологического мониторинга. В таежной зоне Тюменской области за период с 1985 по 2002 год отмечено 4 пика численности и 5 депрессий (Рис. 4), то же самое отмечают Н.С.Москвитина с соавторами (2000) для лесных полевок в Томском Приобье. Причем, флуктуации численности насекомоядных и грызунов, по нашим данным, идут синхронно, хотя, по мнению ряда авторов (Бачурин и др., 1975), эти колебания у разных видов мелких млекопитающих могут не совпадать, и тогда состав доминантов по годам будет меняться. Так летом 2002 года при невысокой численности грызунов отмечалось высокое обилие насекомоядных, обеспечившее некоторый рост численности мелких млекопитающих в целом. Динамика численности мелких млекопитающих в разных биогеоценозах близка, хотя в ряде из них, играющих роль «стаций переживания» (в Среднем Приобье это - спелые зеленомошно-кустарничковые кедровники), численность меняется не так значительно, оставаясь достаточно высокой даже в годы депрессий.
При попытке установить корреляционные зависимости численности красной полевки и обыкновенной бурозубки с различными морфофизиологическими характеристиками в предшествующий вегетационный сезон наиболее сильная положительная связь отмечена с индексом печени самок-сеголеток (r = 0.72 ± 0.40 и 0.91 ± 0.24 при P < 0.05 соответственно для каждого вида). Корреляционной связи обилия названных выше доминирующих в таежной зоне видов с успешностью их размножения или с солнечной активностью (в предыдущий сезон) в 1987-97 гг. нами не установлено. Зная естественные закономерности динамики численности тех или иных видов млекопитающих в определенных регионах, по наблюдаемым отклонениям от них можно выносить суждения о влиянии каких-то возмущающих факторов на популяцию вида, что чрезвычайно важно в системе экологического мониторинга. Примером таких наблюдений являются наши исследования влияния нефтедобычи (в первую очередь, нефтяного загрязнения) на мелких млекопитающих в Среднем Приобье Тюменской области в 1987-1991 гг.
Нужно отметить, что биогеоценотическая роль ряда видов из формального списка минимальна в силу их естественной невысокой плотности популяций (летяга, крошечная бурозубка, тушканчик большой и др.), либо в силу «случайного» характера пребывания на территории Тюменской области (к последним относятся представители категории «возможно встречающихся» видов). Ценность таких видов для целей экологического мониторинга ниже, однако, было бы методологически ошибочно исключать их из поля зрения, так как они могут являться индикаторами изменения например климатических условий существования на границе своего ареала. В фауне млекопитающих Тюменской области установлены и сезонные изменения видового состава, распространения и численности популяций видов. Так, зимой отсутствуют все виды рукокрылых, широко к югу расселяется песец; на севере таежной зоны сокращается численность лося и увеличивается численность северного оленя, а на юге резко снижается количество косули; часть видов (бурый медведь, бурундук, суслики и др.) впадает в спячку, и их роль в биогеоценозе также резко снижается (Лаптев, 1958).
|
Рис. 3. Солнечная активность (А) и динамика численности мелких млекопитающих в разных природных зонах Тюменской области (Б).
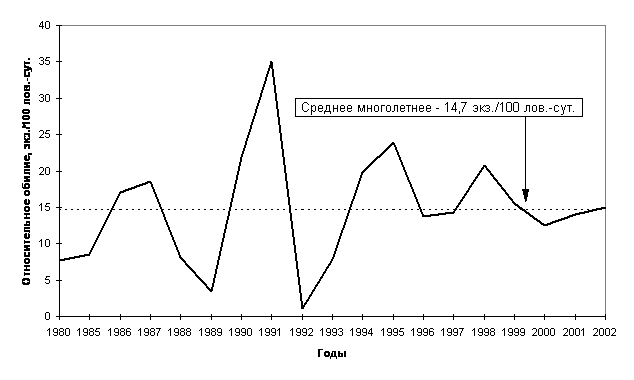
Рис. 4. Динамика численности мелких млекопитающих в таежной зоне
Тюменской области.