Физиологические функции крови
Транспортная функция крови состоит в том, что она переносит газы, питательные вещества, продукты обмена веществ, гормоны, медиаторы, электролиты, ферменты и др
Дыхательная функция заключается в том, что гемоглобин эритроцитов переносит кислород от легких к тканям организма, а углекислый газ от клеток к легким.
Питательная функция — перенос основных питательных веществ от органов пищеварения к тканям организма.
Экскреторная функция (выделительная) осуществляется за счет транспорта конечных продуктов обмена веществ (мочевины, мочевой кислоты и др.) и лишних количеств солей и воды от тканей к местам их выделения (почки, потовые железы, легкие, кишечник).
Водный баланс тканей зависит от концентрации солей и количества белка в крови и тканях, а также от проницаемости сосудистой стенки.
Регуляция температуры тела осуществляется за счет физиологических механизмов, способствующих быстрому перераспределению крови в сосудистом русле. При поступлении крови в капилляры кожи теплоотдача увеличивается, переход же ее в сосуды внутренних органов способствует уменьшению потери тепла.
Защитная функция - кровь является важнейшим фактором иммунитета. Это обусловлено наличием в крови антител, ферментов, специальных белков крови, обладающих бактерицидными свойствами, относящихся к естественным факторам иммунитета.
Одним из важнейших свойств крови является ее способность свертываться, что при травмах предохраняет организм от кровопотери.
Регуляторная функция заключается в том, что поступающие в кровь продукты деятельности желез внутренней секреции, пищеварительные гормоны, соли, ионы водорода и др. через центральную нервную систему и отдельные органы (либо непосредственно, либо рефлекторно) изменяют их деятельность.
2. Химический состав крови. Минеральные и органические вещества.
Периферическая кровь состоит из жидкой части — плазмы и взвешенных в ней форменных элементов.
В состав плазмы крови входят вода (90—92%) и сухой остаток (8—10%). Сухой остаток состоит из органических и неорганических веществ.
К органическим веществам плазмы крови относятся: 1) белки плазмы — альбумины (около 4,5%), глобулины (2—3,5%), фибриноген (0,2—0,4%). Общее количество белка в плазме составляет 7—8%;
2) небелковые азотсодержащие соединения (аминокислоты, полипептиды, мочевина, мочевая кислота, креатин, креатинин, аммиак). Общее количество небелкового азота в плазме (так называемого остаточного азота) составляет 11 —15 ммоль/л (30—40 мг%). При нарушении функции почек, выделяющих шлаки из организма, содержание остаточного азота в крови резко возрастает;
3) безазотистые органические вещества: глюкоза — 4,4—6,65 ммоль/л (80—120 мг%), нейтральные жиры, липиды;
4) ферменты и проферменты: некоторые из них участвуют в процессах свертывания крови и фибринолиза, в частности протромбин и профибринолизин. В плазме содержатся также ферменты, расщепляющие гликоген, жиры, белки и др.
Неорганические вещества плазмы крови составляют около 1 % от ее состава. К этим веществам относятся преимущественно катионы — Ка+, Са2+, К+, Мg2+ и анионы Сl, НРO4, НСО3.
3. Особенности строения, развития и метаболизма эритроцита.
Двояковогнутая форма эритроцитов имеет большую площадь поверхности по сравнению с клетками сферической формы такого же размера. Это облегчает газообмен между клеткой и внеклеточной средой. Кроме того, такая форма, а также особенности строения мембраны и цитоскелета обеспечивают большую пластичность эритроцитов при прохождении ими мелких капилляров. Важную роль в сохранении формы и способности к обратимой деформации эритроцитов играют липиды и белки плазматической мембраны. Липиды бислоя плазматической мембраны эритроцитов, так же, как плазматические мембраны других клеток, содержат глицерофосфолипиды, сфингофосфолипиды, гликолипиды и холестерол.
В процессе дифференцировки эритроцита на стадии эритробласта происходят интенсивный синтез гемоглобина, конденсация хроматина, уменьшение размера ядра и его удаление. Образующийся ретикулоцит ещё содержит глобиновую мРНК и активно синтезирует гемоглобин. Циркулирующие в крови ретикулоциты лишаются рибосом, ЭР, митохондрий и в течение двух суток превращаются в эритроциты. Стволовая клетка превращается в эритроцит за две недели. Эритроциты не содержат ядра и поэтому не способны к самовоспроизведению и репарации возникающих в них повреждений. Эти клетки циркулируют в крови около 120 дней и потом разрушаются макрофагами в печени, селезёнке и костном мозге.
Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками - присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином.
4. Гемоглобин, строение, свойства, биологическая роль.
Гемоглобин является сложным белком, хромопротеидом, состоящим из белка (глобина) и простетической группы — гема. Одна молекула гемоглобина содержит 4 гема. Гем представляет собой комплекс закисного железа с протопорфирином — сложным органическим соединением, принадлежащим очень важному классу веществ — порфиринам.Гемоблобины выполняют 2 важные функции:
- перенос О2 из лёгких к периферическим тканям;
- участие в переносе СО2 и протонов из периферических тканей в лёгкие для последующего выведения из организма.
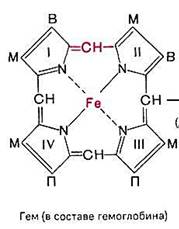 Таким образом, олигомерный белок гемоглобин, в отличие от мономерного родственного белка миоглобина, способен присоединять к специфическим участкам 4 различных лиган-да: О2, Н+, СО2 и БФГ. Все эти лиганды присоединяются к пространственно разобщённым участкам, но конформационные изменения белка в месте присоединения одного лиганда передаются на весь олигомерный белок и изменяют сродство к нему других лигандов. Так, количество поступающего в ткани О2 зависит не только от парциального давления О2, но и концентрации аллостерических лигандов, что увеличивает возможность регуляции функций гемоглобина.
Таким образом, олигомерный белок гемоглобин, в отличие от мономерного родственного белка миоглобина, способен присоединять к специфическим участкам 4 различных лиган-да: О2, Н+, СО2 и БФГ. Все эти лиганды присоединяются к пространственно разобщённым участкам, но конформационные изменения белка в месте присоединения одного лиганда передаются на весь олигомерный белок и изменяют сродство к нему других лигандов. Так, количество поступающего в ткани О2 зависит не только от парциального давления О2, но и концентрации аллостерических лигандов, что увеличивает возможность регуляции функций гемоглобина.
Как мы уже рассматривали выше, в капиллярах работающей мышцы увеличение концентрации СО2 и Н+ уменьшает сродство гемоглобина к О2 и увеличивает отдачу его в ткани. При длительной гипоксии усиливается синтез 2,3-БФГ в эритроцитах, что также снижает сродство гемоглобина к О2 и при том же парциальном давлении О2 увеличивает его транспорт в ткани.
Следовательно, благодаря воздействию регуляторных лигандов олигомерные белки способны приспосабливать свою конформацию и фунцию к изменениям, происходящим в окружающей среде.
5. Варианты первичной структуры гемоглобина человека. Гемоглобинопатии.