Тип простейших (Protozoa) состоитиз 5 классов: Саркодовые (Sarcodina), Жгутиконосцы (Mastigophora), Споровики (Sporozoa), Книдоспоридии(Cnidosporidia) и Инфузории (Infusoria)
Общая характеристика
Первый тип, с которого мы начинаем знакомство с миром животных,- это тип простейших (Protozoa). Он состоит из многих классов, отрядов, семейств и включает примерно 20-25 тыс. видов.
Простейшие распространены на всей поверхности нашей планеты и живут в самых различных средах. В большом количестве мы найдем их в морях и океанах, как непосредственно в толще морской воды, так и на дне. Обильны простейшие в пресных водах. Некоторые виды живут в почве.
Многие простейшие в качестве среды обитания используют другие организмы, ведут паразитический образ жизни. Среди паразитических простейших имеются возбудители тяжелых заболеваний человека, домашних и промысловых животных.
По своему строению простейшие чрезвычайно разнообразны. Подавляющее большинство их обладает микроскопически малыми размерами, для изучения их приходится пользоваться микроскопом.
Основной и самой характерной чертой простейших является их одноклеточность. Простейшие являются организмами, тело которых по строению соответствует одной клетке.
Если по своему строению простейшие соответствуют клеткам многоклеточных организмов, то в функциональном отношении они несравнимы с ними. Клетка в теле многоклеточного всегда представляет собой только часть организма, ее отправления подчинены функциям многоклеточного организма как целого. Напротив, простейшее - это самостоятельный организм, которому свойственны все жизненные функции: обмен веществ, раздражимость, движение, размножение.
К окружающим условиям внешней среды простейшее приспосабливается как целый организм. Следовательно, можно сказать, что простейшее - это самостоятельный организм на клеточном уровне организации.
Размеры тела подавляющего большинства простейших микроскопические. Наиболее мелкие представители типа имеют 2-4 микрона* в поперечнике (например, паразитические жгутиконосцы из рода лейшманий - Leichmania, разные виды семейства пироплазмид, паразитирующие в красных кровяных клетках млекопитающих).
Наиболее обычные размеры простейших - в пределах 50-150 мк. Но среди них имеются и гораздо более крупные организмы.
Инфузории Bursaria, Spirostomum достигают 1,5 мм длины - их хорошо видно простым глазом, грегарины Porospora gigantea - длины до 1 см.
У некоторых корненожек фораминифер раковина достигает 5-6 см в диаметре (например, виды рода Psammonix, ископаемые нуммулиты и др.).
Низшие представители простейших (например, амебы) не обладают постоянной формой тела. Их полужидкая цитоплазма постоянно меняет свои очертания благодаря образованию разнообразных выростов - ложных ножек (cм. рис. 24), служащих для движения и захвата пищи.
Большинство же простейших обладает относительно постоянной формой тела, которая обусловлена наличием опорных структур. Среди них наиболее обычной является плотная эластичная мембрана (оболочка), образуемая периферическим слоем цитоплазмы (эктоплазмой) и носящая название пелликулы.
В одних случаях пелликула относительно тонка и не препятствует некоторому изменению формы тела простейшего, как это имеет место, например, у способных сокращаться инфузорий. У других простейших она образует прочный и не меняющий своей формы наружный панцирь.
У многих жгутиконосцев, окрашенных в зеленый цвет благодаря наличию хлорофилла, имеется наружная оболочка из клетчатки - признак, характерный для растительных клеток.
Что касается общего плана строения и элементов симметрии, то простейшие обнаруживают большое разнообразие. Такие животные, как амебы, не обладающие постоянной формой тела, не имеют постоянных элементов симметрии.
Широко распространены среди Protozoa разные формы радиальной симметрии, свойственной главным образом планктонным формам (многиерадиолярии, солнечники.). При этом имеется один центр симметрии, от которого отходит различное число пересекающихся в центре осей симметрии, определяющих расположение частей тела простейшего.
У многих радиально построенных форм можно выделить одну главную ось, определяющую передний и задний концы тела, вокруг которой радиально располагаются части тела простейшего (некоторые радиолярии, см. табл. 2, 3, инфузории Didinium.).
Относительно редко встречается у простейших двубоковая (билатеральная) симметрия, при которой можно провести одну-единственную* плоскость симметрии, делящую тело животного на две равные зеркальные половины (раковины некоторых фораминифер, см. рис. 32, 33,радиолярии, см. табл. 2 и 3, некоторые виды жгутиконосцев, например лямблия, см. далее рис. 57). Большинство простейших из разных классов являются асимметричными.
У сложно организованных простейших из класса инфузорий и у некоторых жгутиконосцев, кроме пелликулы, имеются еще и другие опорные структуры, поддерживающие и определяющие форму тела. К ним относятся тончайшие волоконца (фибриллы), проходящие в различных направлениях. Примером могут служить опорные волоконца одной из инфузорий.
На рисунке 19 видно, какой большой сложности может достигать эта система, образующая прочный и эластичный каркас, поддерживающий полужидкую цитоплазму простейшего.

Рис. 19. Опорные (скелетные) фибриллы и ресничный аппарат круго-ресничной инфузории Trichodina
К числу опорных и вместе с тем защитных образований у простейших относятся различные формы минерального скелета, свойственного преимущественно многим представителям класса саркодовых. Эти скелетные образования чаще всего имеют форму раковинок, иногда очень сложно устроенных (в отряде фораминифер). В других случаях основу скелета составляют отдельные иглы (спикулы), обычно соединяющиеся между собой. По химическому составу минеральный скелет простейших различен. Наиболее обычными компонентами его являются углекислый кальций (СаСO3) или окись кремния (SiO2). Более подробно строение скелета будет рассмотрено при знакомстве с отдельными классами простейших.
Большинству простейших, за исключением некоторых паразитических форм, свойственна способность к движению - перемещению в пространстве. Формы движения простейших разнообразны. Наиболее простой и, вероятно, исходной формой является амебоидное движение. Оно выражается в образовании ложных ножек (псевдоподий) - выростов цитоплазмы разной формы. Все содержимое клетки как бы медленно перетекает в направлении образующейся псевдоподии, и таким путем осуществляется перемещение простейшего в пространстве. Эта форма движения преимущественно свойственна представителям класса саркодовых. Разным видам свойственна различная форма псевдоподий (см. далее рис. 26).
Более сложной формой является движение, осуществляемое при помощи жгутиков и ресничек. Жгутиковая форма движения характерна длякласса жгутиконосцев.
Жгутики представляют собой тончайшие выросты тела. Количество их у разных видов различно - от одного до многих десятков и даже сотен (см. далее рис. 40, 63). Каждый жгутик берет начало от небольшого базального зернышка, называемого блефаропластом и расположенного в цитоплазме. Таким образом, непосредственно граничащая с базальным зерном часть жгутика проходит внутри цитоплазмы (она носит названиекорневой нити), а затем проходит через пелликулу наружу. Механизм жгутикового движения у разных видов различен. В большинстве случаев он сводится к вращательному движению. Жгутик описывает фигуру конуса, вершиной обращенного к месту его прикрепления. Наибольший механический эффект достигается, когда угол, образуемый вершиной конуса, составляет 40-46°. Быстрота движения различна, она колеблется у разных видов между 10 и 40 оборотами в секунду. Простейшее как бы "ввинчивается" в окружающую его жидкую среду.
Нередко вращательное движение жгутика сочетается с его волнообразным движением. Обычно при поступательном движении само тело простейшего вращается вокруг продольной оси.
Изложенная схема справедлива для большинства одножгутиковых форм. У многожгутиковых движение жгутиков может носить иной характер, в частности жгутики могут находиться в одной плоскости, не образуя конуса вращения.
Электронномикроскопические исследования последних лет показали, что внутренняя ультрамикроскопическая структура жгутиков весьма сложна. Снаружи жгутик окружен тонкой мембраной, которая является непосредственным продолжением самого поверхностного слоя эктоплазмы - пелликулы. Внутренняя полость жгутика заполнена цитоплазматическим содержимым. По продольной оси жгутика проходит одиннадцать тончайших нитей (фибрилл), которые нередко являются двойными (рис. 20). Эти фибриллы располагаются всегда закономерно. Девять из них (простых или двойных) лежат по периферии, образуя в совокупности как бы цилиндр. Две фибриллы занимают центральное положение. Чтобы составить себе представление о размерах всех этих образований, достаточно сказать, что диаметр периферических фибрилл составляет около 350Å (ангстрем). Ангстрем-единица длины, равная 0,0001 дк, а микрон равняется 0,001 мм. Вот какие ничтожные по своим размерам структуры стали доступными для изучения благодаря внедрению в микроскопическую технику электронного микроскопа.
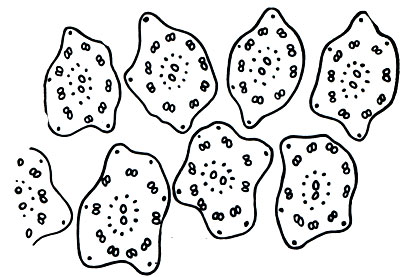
Рис. 20. Поперечный разрез жгутиков Pseudotrichonympha (рисунок сделан с электронно-микроскопической фотографии). Видны в каждом жгутике девять периферических двойных фибрилл и центральная пара фибрилл
Функциональное значение фибрилл жгутиков не может считаться окончательно выясненным. По-видимому, часть их (вероятно, периферические) играет активную роль в двигательной функции жгутика п содержит особые белковые молекулы, способные сокращаться, другие же являются опорными эластическими структурами, имеющими поддерживающее значение.
Реснички служат органоидами движения инфузорий. Обычно число их у каждой особи очень велико и измеряется несколькими сотнями, тысячами и даже десятками тысяч. Механизм движения ресничек несколько иной, чем жгутиков. Каждая ресничка совершает гребные движения. Она быстро и с силой сгибается в одну сторону, а затем медленно выпрямляется.
Совместное действие большого числа ресничек, биение которых координировано, вызывает быстрое поступательное движение простейшего.
Каждая ресничка инфузории, как показали новейшие исследования, является сложным образованием, по своему строению соответствующим жгутику. У основания каждой реснички всегда располагается так называемое базальное зерно (иначе, кинетозома) - важная часть ресничного аппарата.
У многих инфузорий отдельные реснички соединяются друг с другом, образуя структуры более сложного строения (мембранеллы, цирры и др.) и более эффективного механического действия.
Некоторым высокоорганизованным простейшим (инфузориям, радиоляриям) свойственна еще одна форма движения - сокращение. Тело таких простейших способно быстро менять свою форму, а затем вновь возвращаться к исходному состоянию.
Способность к быстрому сокращению обусловлена наличием в теле простейшего особых волоконец - мионем - образований, аналогичных мышцам многоклеточных животных.
У некоторых простейших существуют еще и другие формы движения.
По способам и характеру питания, по типу обмена веществ простейшие обнаруживают большое разнообразие.
В классежгутиконосцев имеются организмы, способные подобно зеленым растениям при участии зеленого пигмента хлорофилла усваивать неорганические вещества - углекислый газ и воду, превращая их в органические соединения (аутотрофный тип обмена). Этот процесс фотосинтеза протекает с поглощением энергии. Источником последней является лучистая энергия - солнечный луч.
Таким образом, эти простейшие организмы правильнее всего рассматривать как одноклеточные водоросли. Но наряду сними в пределах того же класса жгутиконосцев имеются бесцветные (лишенные хлорофилла) организмы, неспособные к фотосинтезу и обладающие гетеротрофным(животным) типом обмена веществ, т. е. питающиеся за счет готовых органических веществ. Способы животного питания простейших, так же как и характер пищи их, очень разнообразны. Наиболее просто устроенные простейшие не обладают специальными органоидами захвата пищи. У амеб, например, псевдоподии служат не только для движения, но вместе с тем и для захвата оформленных частиц пищи. У инфузорий для захвата пищи служит ротовое отверстие. С последним обычно связаны разнообразные структуры - околоротовые мерцательные перепонки (мембранеллы), способствующие направлению пищевых частиц к ротовому отверстию и далее в особую трубку, ведущую в эндоплазму - клеточную глотку.
Пища простейших очень разнообразна. Одни питаются мельчайшими организмами, например бактериями, другие - одноклеточными водорослями, некоторые являются хищниками, пожирающими других простейших, и т. п. Непереваренные остатки пищи выбрасываются наружу - у саркодовых на любом участке тела, у инфузорий через особое отверстие в пелликуле.
У пресноводных и у части морских простейших имеется особый органоид, связанный с регуляцией осмотического давления в клетке простейшего и с выделением. Это сократительная вакуоля. Она представляет собой периодически появляющийся в цитоплазме пузырек, наполняющийся жидкостью и опорожняющийся наружу. Сократительная вакуоля регулирует количество воды в цитоплазме, поступающей из окружающей среды благодаря разности осмотических давлений. У паразитических простейших и у многих морских форм, живущих в среде с повышенным осмотическим давлением, сократительные вакуоли отсутствуют.
Особых органоидов дыхания у простейших нет, они поглощают кислород и выделяют углекислоту всей поверхностью тела.
Как и все живые существа, простейшие обладают раздражимостью, т.е. способностью отвечать той или иной реакцией на факторы, действующие извне. Простейшие реагируют на механические, химические, термические, световые, электрические и иные раздражения. Реакции простейших на внешние раздражения часто выражаются в изменении направления движения и носят название таксис. Таксисы могут быть положительными, если движение осуществляется в направлении раздражителя, и отрицательными, если оно осуществляется в противоположную сторону.
Реакции многоклеточных животных на раздражения осуществляются под воздействием нервной системы. Многие исследователи пытались обнаружить и у простейших (т. е. в пределах клетки) аналоги нервной системы. Американские ученые, например, описывали у многих инфузорий наличие особого нервного центра (так называемого моториума), представляющего собой особый уплотненный участок цитоплазмы. От этого центра к различным участкам тела инфузории отходит система тонких волоконец, которые рассматривались как проводники нервных импульсов. Другие исследователи, применяя особые методы серебрения препаратов (обработка азотнокислым серебром с последующим восстановлением металлического серебра), обнаружили в эктоплазме инфузорий сеть тончайших волоконец. Эти структуры (рис. 21) также рассматривались как нервные элементы, по которым распространяется волна возбуждения. В настоящее время, однако, большинство ученых, изучающих тонкие фибриллярные структуры, придерживаются иного мнения об их функциональной роли в клетке простейшего. Экспериментальных доказательств нервной роли фибриллярных структур не получено. Напротив, имеются опытные данные, которые дают возможность предполагать, что у простейших волна возбуждения распространяется непосредственно по наружному слою цитоплазмы - эктоплазме. Что же касается различного рода фибриллярных структур, которые еще недавно рассматривались как "нервная система" простейших, то они имеют скорее всего опорное (скелетное) значение и способствуют сохранению формы тела простейшего.

Рис. 21. Инфузория Urocentrum turbo. Назальные зерна ресничек и система эктоплазматических волоконец (фибрилл), выявляемые методом импрегнации серебром
Как и всякая клетка, простейшие имеют ядро. Выше, при рассмотрении строения клетки, мы уже ознакомились с основными структурными компонентами ядра. В ядрах простейших, так же как и в ядрах многоклеточных, имеется оболочка, ядерный сок (кариолимфа), хроматин(хромосомы) и ядрышки. Однако по размерам и строению ядра разные простейшие весьма разнообразны (рис. 22). Эти различия обусловлены соотношением структурных компонентов ядра: количеством ядерного сока, количеством и размерами ядрышек (нуклеол), степенью сохранения строения хромосом в интерфазном ядре и т. п.

Рис. 22. Ядра различных простейших: А - микронуклеус инфузории Paramecium aurelia; Б - жгутиконосец Trypanosoma brucei; В - амеба Amoeba sphaeronuclei; Г - панцирный жгутиконосец Ceratium fuscus; Д - радиолярия Aulacantha scolymantha; E - макрогамета кокцидии Aggregata eberthi; Ж - макронуклеус инфузории Epidinium ecaudatum
У большинства простейших имеется одно ядро. Однако встречаются и многоядерные виды простейших.
У некоторых простейших, а именно у инфузорий и немногих корненожек - фораминифер, наблюдается интересное явление дуализма(двойственности) ядерного аппарата. Оно сводится к тому, что в теле простейшего имеются два ядра двух категорий, различающиеся как по своему строению, так и по физиологической роли в клетке. У инфузорий, например, имеется два типа ядер: большое, богатое хроматином ядро -макронуклеус и маленькое ядро - микронуклеус. Первое связано с выполнением вегетативных функций в клетке, второе-с половым процессом.
Простейшим, как и всем организмам, свойственно размножение. Существуют две основные формы размножения простейших: бесполое и половое. В основе того и другого лежит процесс деления клетки.
При бесполом размножении число особей возрастает в результате деления. Например, амеба при бесполом размножении делится на две амебы путем перетяжки тела. Процесс этот начинается с ядра, а затем захватывает цитоплазму. Иногда бесполое размножение приобретает характер множественного деления. При этом ядро предварительно делится несколько раз и простейшее становится многоядерным. Вслед за этим цитоплазма распадается на число отдельностей, соответствующих количеству ядер. В результате организм простейшего сразу дает начало значительному количеству мелких особей. Так происходит, например, бесполое размножение малярийного плазмодия - возбудителя малярии человека.
Половое размножение простейших характеризуется тем, что собственно размножению (увеличению числа особей) предшествует половой процесс, характерным признаком которого является слияние двух половых клеток (гамет) или двух половых ядер, ведущее к образованию одной клетки - зиготы, дающей начало новому поколению. Формы полового процесса и полового размножения у простейших в высшей степени разнообразны. Основные формы его будут рассмотрены при изучении отдельных классов.
Многие виды простейших имеют не одну, а несколько форм размножения, которые могут закономерно чередоваться друг с другом. В результате получается сложный цикл развития, отдельные этапы которого могут протекать в разных условиях среды. Особенной сложностью отличаются циклы развития у паразитических простейших в классеспоровиков.
Простейшие обитают в самых различных условиях среды. Большинство их - водные организмы, широко распространенные как в пресных, так и в морских водоемах. Многие виды их живут в придонных слоях и входят в состав бентоса. Большой интерес представляет приспособление простейших к жизни в толще песка, в толще воды (планктон).
Небольшое число видов Protozoa приспособилось к жизни в почве. Их средой обитания являются тончайшие пленки воды, окружающие почвенные частицы и заполняющие капиллярные просветы в почве. Интересно отметить, что даже в песках пустыни Каракум живут простейшие. Дело в том, что под самым верхним слоем песка здесь расположен влажный слой, пропитанный водой, приближающейся по своему составу к морской воде. В этом влажном слое и были обнаружены живые простейшие из отряда фораминифер, являющиеся, по-видимому, остатками морской фауны, населявшей моря, ранее находившиеся на месте современной пустыни. Эта своеобразная реликтовая фауна в песках Каракумов впервые была обнаружена проф. Л. Л. Бродским при изучении воды, взятой из колодцев пустыни.
Очень многие простейшие, относящиеся к различным классам этого типа, ведут паразитический образ жизни. Их среда обитания и источники пищи - другие живые организмы. Свыше 3,5 тыс. видов простейших - паразиты. Хозяевами их являются самые различные животные и растения. Многие паразитические простейшие живут в организме человека, домашних и промысловых животных, нанося им большой вред. Один из классов типа простейших - споровики - целиком состоит из паразитов.
Практическое значение простейших для человека велико. В особенности это относится к паразитам. До настоящего времени в тропических зонах земного шара широко распространена малярия - тяжелое заболевание, поражающее ежегодно десятки миллионов человек в Индии и других тропических частях Азии, в Африке и других странах. Возбудитель этого заболевания относится к классу споровиков типа простейших. Тяжелым заболеванием человека в Центральной Африке является сонная болезнь, вызываемая паразитом из класса жгутиконосцев. Большой ущерб наносят животноводству заболевания домашних животных, вызываемые простейшими. Сюда относятся различные пироплазмозы, кокцидиозы, трипанозомозы и многие другие. Важнейшие из них в практическом отношении мы рассмотрим ниже.
Имеется ряд простейших паразитов из отрядакокцидий, поражающих домашнюю птицу, в особенности кур. Борьба с этими многочисленными и опасными протозойными заболеваниями требует детального изучения биологии возбудителей, их циклов развития.
Некоторый практический интерес представляют и свободноживущие простейшие. Разные виды их приурочены к определенному комплексу внешних условий, в частности к различному химическому составу воды.
Определенные виды простейших живут при разной степени загрязненности пресных вод органическими веществами. Поэтому по видовому составу простейших можно судить о свойствах воды водоема. Эти особенности простейших используют для санитарно-гигиенических целей при так называемом биологическом анализе воды.
В общем круговороте веществ в природе простейшие играют заметную роль. В водоемах многие из них являются энергичными пожирателями бактерий и других микроорганизмов. Вместе с тем сами они служат пищей для более крупных животных организмов. В частности, выклевывающиеся из икринок мальки многих видов рыб на самых начальных этапах своей жизни питаются преимущественно простейшими.
Типпростейших в геологическом отношении является весьма древним. В ископаемом состоянии хорошо сохранились те виды простейших, которые обладали минеральным скелетом (фораминиферы, радиолярии). Ископаемые остатки их известны начиная с самых древних нижнекембрийских отложений.
Морские простейшие - корненожки и радиолярии - играли и играют весьма существенную роль в образовании морских осадочных пород. В течение многих миллионов и десятков миллионов лет микроскопически мелкие минеральные скелеты простейших после отмирания животных опускались на дно, образуя здесь мощные морские отложения. При изменении рельефа земной коры, при горнообразовательных процессах в прошлые геологические эпохи, морское дно становилось сушей. Морские осадки превращались в осадочные горные породы. Многие из них, как, например, некоторые известняки, меловые отложения и др., в значительной своей части состоят из остатков скелетов морских простейших. В силу этого изучение палеонтологических остатков простейших играет большую роль в определении возраста разных слоев земной коры и, следовательно, имеет существенное значение при геологической разведке, в частности при разведке полезных ископаемых.
Тип простейших (Protozoa) состоитиз 5 классов: Саркодовые (Sarcodina), Жгутиконосцы (Mastigophora), Споровики (Sporozoa), Книдоспоридии(Cnidosporidia) и Инфузории (Infusoria).
Класс саркодовые (Sarcodina)
К классу саркодовых относятся наиболее просто устроенные среди простейших организмы. Как уже было сказано в общем очерке организации типа простейших, в подразделении его на классы существенную роль играют органоиды движения. Для саркодовых характерной формой их являются ложноножки, или псевдоподии, которые представляют собой временно образующиеся выросты цитоплазмы. Псевдоподии служат вместе с тем для захвата пищи.
Строение саркодовых, несмотря на относительную простоту их организации, отличается большим разнообразием. Главным образом это касается скелетных образований, которые, как увидим дальше, достигают у саркодовых большой сложности и совершенства.
Общее количество видов живущих в настоящее время саркодовых измеряется числом 8-10 тыс. Очень большое количество видов известно в ископаемом состоянии благодаря хорошей сохранности скелетов многих групп саркодовых.
Свыше 80% всех современных саркодовых являются обитателями моря. Некоторые виды живут в пресной воде, немногие приспособились к жизни в почве. Имеются также паразитические виды.
Класссаркодовых состоит из трех подклассов, весьма неравноценных по количеству относящихся к ним видов:
1. Корненожки (Rhizopoda) - около 2 тыс. видов;
2. Солнечники (Helizoa) -всего несколько десятков видов;
3. Лучевики (Radiolaria) - 7-8 тыс. видов.
Рассмотрим наиболее интересных и характерных представителей этих трех подклассов.
Подкласс корненожки (Rhizopoda)
Наиболее просто устроенными организмами среди корненожек являются голые амебы (Amoebina), образующие первый отряд подкласса корненожек.
Чтобы познакомиться со строением и образом жизни голых амеб, рассмотрим сначала какого-нибудь одного характерного и часто встречающегося представителя.
Амеба протей (Amoeba proteus). В пресных водах, в небольших прудах и канавах с илистым дном, нередко удается обнаружить амебу протея (Amoeba proteus). Культуру этого вида легко развести в лабораторных условиях. Амеба протей - одна из крупных свободноживущих амеб. В активном состоянии она достигает размера 0,5 мм, ее видно простым глазом. Если наблюдать под микроскопом за живой амебой (рис. 23, 24), видно, что она образует несколько довольно длинных лопастных, тупо заканчивающихся псевдоподий. Псевдоподии все время меняют свою форму, часть их втягивается внутрь, часть, напротив, удлиняется, иногда разветвляется. Тело амебы как бы переливается в псевдоподии, которые в нескольких точках прикрепляются к субстрату, и благодаря этому образование ложных ножек приводит к поступательному движению всей амебы. Псевдоподии служат не только для движения, но и для заглатывания пищи. Если псевдоподия в процессе своего образования наталкивается на какую-либо органическую частицу (водоросль, мелкое простейшее и т. п.), она "обтекает" ее со всех сторон (рис. 25) и включает внутрь цитоплазмы вместе с небольшим количеством жидкости.

Рис. 23. Амебы в капле воды (Amoeba proteus) (вверху)

Рис. 24. Амеба протей (Amoeba proteus): 1 - эктоплазма; 2 - эндоплазма; 3 - непереваренные частицы пищи, выбрасываемые наружу; 4 - сократительная вакуоля; 5 - ядро; 6 - пищеварительная: вакуоля (внизу)

Рис. 25. Последовательные стадии заглатывания пищи амебой (Amoeba terricola)
Таким образом в цитоплазме образуются пузырьки с пищевыми включениями, которые называют пищеварительными вакуолями. В них происходит переваривание пищи (внутриклеточное пищеварение).
Не переваренные остатки пищи через некоторое время выбрасываются наружу (см. рис. 24).
Вся цитоплазма амебы ясно подразделена на два слоя. Наружный, светлый, вязкий, всегда лишенный пищеварительных вакуолей, носит название эктоплазмы. Внутренний, зернистый, гораздо более жидкий, несущий многочисленные пищевые включения, называют эндоплазмой. В состав псевдоподий входят оба слоя цитоплазмы. Эктоплазма и эндоплазма не представляют собой резко разграниченных частей тела амебы. Они могут превращаться друг в друга. В области образования и нарастания псевдоподии, куда устремляется жидкая эндоплазма, периферические части ее желатинизируются (уплотняются) и превращаются в эктоплазму.
Напротив, на противоположном конце тела протекает обратный процесс - разжижение эктоплазмы и частичное превращение ее в эндоплазму. Это явление обратимого превращения эндоплазмы в эктоплазму и обратно лежит в основе образования псевдоподий.
Кроме пищевых включений (часто сосредоточенных в пищеварительных вакуолях), в цитоплазме амебы протея обычно отчетливо бывает виден светлый пузырек, который периодически то появляется, то исчезает. Это сократительная вакуоля, играющая очень важную роль в жизненных отправлениях амебы. Сократительная вакуоля заполняется жидкостью (в основном водой), которая поступает в нее из окружающей цитоплазмы. Достигнув определенного, характерного для данного вида амеб размера, сократительная вакуоля сокращается. Ее содержимое при этом изливается наружу через пору. Весь период наполнения и сокращения вакуоли при комнатной температуре длится у амебы протея обычно 5-8 минут.
Концентрация различных растворенных органических и неорганических веществ в теле амебы выше, чем в окружающей пресной воде. Поэтому в силу законов осмоса вода проникает в протоплазму амебы. Если бы избыток ее не выводился наружу, то через короткий промежуток времени амеба "расползлась" бы и растворилась в окружающей воде. Благодаря деятельности сократительной вакуоли этого не происходит. Таким образом, сократительная вакуоля - это, прежде всего-органоид осморегуляции, регулирующий постоянно осуществляемый ток воды через тело простейшего. Однако наряду с этим она связана и с другими жизненными функциями. Вместе с выводимой из тела амебы жидкостью выводятся и продукты обмена веществ. Следовательно, сократительная вакуоля участвует в функции выделения.
Постоянно поступающая в цитоплазму вода содержит кислород. Поэтому сократительная вакуоль косвенно участвует и в функции дыхания.
Как и во всякой клетке, в теле амебы есть ядро. На живом объекте оно почти не видно. Для выявления ядра применяют некоторые красители, избирательно окрашивающие нуклеиновые вещества ядра. У амебы протея ядро довольно крупное, расположено в эндоплазме, примерно в центре тела.
Как размножаются амебы? Единственной известной у них формой размножения является деление надвое в свободноподвижном состоянии. Процесс этот начинается скариокинетического деления ядра. Вслед за тем на теле амебы появляется перетяжка, которая в конце концов перешнуровывает тело ее на две равные половинки, в каждую из которых отходит по одному ядру. Темп размножения амебы протея зависит от условий, и прежде всего от питания и температуры. При обильном питании и температуре 20-25° С амеба делится один раз в течение 1 - 2 суток.
В пресной и морской воде живет несколько десятков видов амеб. Они различаются размерами, формой псевдоподий (рис. 26). Ложные ножки могут сильно отличаться по форме и размерам. Есть виды амеб (см. рис. 26), у которых образуется всего одна толстая короткая псевдоподия, у других - несколько длинных заостренных, у третьих - много коротких тупых и т. п. Следует отметить, что даже в пределах одного вида амеб форма псевдоподий может довольно широко варьировать в зависимости от условий окружающей среды (солевой состав, кислотность среды и т. п.).

Рис. 26. Разные виды амеб с различной формой псевдоподий: 1 - Amoeba limax; 2 - Pelomyxa binucieata; 3 - Amoeba proteus; 4 - Amoeba radiosa; 5 -Amoeba verrucosa; 6 - Amoeba polypodia (вверху)
Паразитические амебы. Некоторые виды амеб приспособились к паразитическому образу жизни в кишечнике позвоночных и беспозвоночных животных. В толстых кишках человека живет пять видов паразитических амеб. Четыре вида их являются безобидными "квартирантами". Они питаются бактериями, которые в огромном количестве населяют толстую и слепую кишку человека (так же как и всех позвоночных животных), и не оказывают никакого влияния на хозяина. Но один из видов паразитирующих в кишечнике человека амеб - дизентерийная амеба (Entamoeba histolytica) - при определенных условиях может вызвать у человека тяжелое заболевание - особую форму кровавого поноса (колита), болезни, носящей название амебиаза.
Что же представляет собой дизентерийная амеба человека, почему она вызывает заболевание, как она проникает в кишечник?
Дизентерийные амебы живут в толстом кишечнике человека. Это очень мелкие (по сравнению, например, с только что описанной амебой протеем) простейшие. Размеры их - 20-30 мк. При изучении живой амебы под микроскопом ясно видно, что у нее резко разграничены эктои эндоплазма, причем зона эктоплазмы относительно широка (рис. 27). Дизентерийная амеба характеризуется очень активной подвижностью. Она образует немногочисленные короткие широкие псевдоподии, в формировании которых принимает участие почти исключительно эктоплазма.

Рис. 27. Дизентерийная амеба (Entamoeba histolytica), рисунки с живого объекта в различных стадиях движения: 1 - эктоплазма; 2 - эндоплазма; 3 - ядро (внизу)
Дизентерийная амеба широко распространена по всему земному шару. В зависимости от географического положения процент зараженности людей этим паразитом варьирует в среднем от 10 до 30. Но заболевание амебиазом встречается очень редко и приурочено преимущественно к субтропическим и тропическим районам земного шара.
В умеренных и северных широтах в подавляющем большинстве случаев дело ограничивается носительством и клинически выраженные формы амебиаза представляют большую редкость.
Почему же имеется такое расхождение между частотой встречаемости паразита и частотой вызываемого им заболевания? Оказывается, дело в том, что далеко не всегда наличие дизентерийной амебы в кишечнике человека сопровождается болезненными явлениями. В большинстве случаев амеба не приносит своему хозяину-человеку никакого вреда. Она живет в просвете кишечника, активно двигается и питается бактериями. Это явление, когда возбудитель какого-либо заболевания присутствует в организме хозяина, но не вызывает патологических явлений, носит название носительства. В отношении дизентерийной амебы имеет место именно носительство.
Иногда амеба меняет свое поведение. Она активно внедряется в стенки кишечника, разрушает эпителий, выстилающий кишку, и проникает в соединительную ткань. Происходит изъязвление стенки кишечника, которое приводит к тяжелой форме кровавого поноса. Амебы, проникшие в ткани, меняют и характер своего питания. Вместо бактерий они начинают активно пожирать красные кровяные клетки (эритроциты). В цитоплазме амеб скапливается большое количество эритроцитов на разных стадиях переваривания (рис. 28). Медицине в настоящее время известны некоторые специфические лекарственные вещества, применение которых убивает амеб, что приводит к выздоровлению. Если не прибегать к лечениюг то амебиаз переходит в хроническую форму и, вызывая тяжелое истощение организма человека, иногда приводит к смертельному исходу.

Рис. 28. Дизентерийная амеба (Entamoeba histolytica): А - амеба с заглоченными красными кровяными клетками; В - амеба без эритроцитов. 1 - ядро; 2 - эритроциты
До сих пор остаются неизвестными причины, которые превращают безобидного "квартиранта" кишечника в "агрессивного" пожирателя тканей. Высказывалось предположение, что существуют разные формы дизентерийной амебы, не отличающиеся друг от друга по своему строению.
Одни из них, распространенные в умеренном и северном поясе, редко переходят к паразитизму в тканях и почти всегда питаются бактериями. Другие - южные - относительно легко становятся "агрессивными" пожирателями тканей.
Каким образом дизентерийная и другие амебы, паразитирующие в кишечнике человека, попадают в организм хозяина?
Активно подвижные формы амеб могут жить только в кишечнике человека. Будучи выведены из него, например в воду, в почву, они погибают очень быстро и не могут служить источником заражения. Заражение осуществляется особыми формами существования амеб - цистами. Посмотрим, как происходит у дизентерийной амебы процесс формирования цист. Попадая вместе с содержимым толстого кишечника в его нижние отделы и в прямую кишку, амебы претерпевают значительные изменения. Они втягивают псевдоподии, выбрасывают пищевые частицы, округляются. Затем эктоплазма выделяет тонкую, но весьма прочную оболочку. Этот процесс представляет собой инцистирование.
Одновременно с выделением оболочки цисты претерпевает изменение и ядро. Оно дважды последовательно делится, причем деление ядра не сопровождается делением цитоплазмы. Таким образом образуются столь характерные для дизентерийной амебы четырех ядерные цисты (рис. 29).

Рис. 29. Стадии инцистирования дизентерийной амебы (Entamoeba histolytica): А - одноядерная предцистная форма; Б - че-тырехъядерная циста. 1 - ядра
В таком виде вместе с фекальными массами цисты выводятся наружу. В отличие от активно подвижных вегетативных форм цисты обладают большой стойкостью. Попадая в воду или в почву, они долгое время сохраняют жизнеспособность (до 2-3 месяцев).
Подсыхание и нагревание гибельны для цист. Доказано, что цисты могут распространяться мухами, сохраняя при этом жизнеспособность.
Попадая в кишечник человека с пищей, водой и т. п., амеба эксцистируется. Ее наружная оболочка растворяется, после чего следуют два деления, не сопровождающиеся делением ядра (цисты, как мы видим, четырехъядерны). В результате получаются четыре одноядерные амебы, которые переходят к активной жизни.
Другие, не патогенные виды амеб кишечника человека распространяются таким же путем при помощи цист. По своему строению (размеры, число ядер) цисты разных видов несколько отличаются друг от друга. На этом основывается их диагностика.
Раковинные корненожки. Кроме амеб, в пресных водах встречаются представители и другого отряда корненожек - раковинные корненожки (Testacea). В море они не встречаются.
По своему строению раковинные корненожки напоминают амеб. В отличие от них часть протоплазматического тела корненожек заключена внутри раковинки, играющей роль защитного образования. В раковинке есть отверстие (устье), через которое наружу выдаются псевдоподии. У арцеллы (Arcella, рис. 30) раковинка имеет форму блюдечка. Устье ее расположено в центре. Раковинка, часто коричневой окраски, состоит из органического вещества, напоминающего по консистенции рог. Выделяется она веществом цитоплазмы подобно тому, как выделяется оболочка цисты. У диффлюгии (Difflugia, рис. 30) раковинка грушевидная. Она состоит из песчинок - мелких посторонних частичек, заглоченных, а затем отложенных на поверхности тела. У эуглифы (Euglypha) раковинка башневидная (рис. 30), но, в отличие от диффлюгии, она слагается из кремневых пластиночек правильной овальной формы. Эти пластиночки образуются в толще цитоплазмы корненожек, а затем выделяются на поверхность. Размеры раковинных корненожек невелики. Обычно они варьируют в пределах 50-150 мк.

Рис. 30. Разные виды раковинных корненожек:: А - Arcella; Б - Difflugia; В - Euglypha - раковинка; Г - Euglypha с псевдоподиями. 1 - псевдоподии; 2 - ядро
Выдающиеся из устья наружу псевдоподии выполняют двоякую функцию. Они служат органоидами движения и захвата пищи. Последнее осуществляется по тому же типу, как и у голых амеб.
В связи с наличием раковины несколько видоизменяется, по сравнению с амебами, способ бесполого размножения - деления. Раковинка служит прочным скелетным образованием, и понятно, что она не может перешнуроваться пополам. Поэтому процесс деления раковинных корненожек связан с развитием новой раковины. Обычно он осуществляется следующим образом. Сначала примерно половина цитоплазмы выступает из устья. Вокруг этой части образуется новая раковинка. Одновременно с этим процессом делится ядро и одно из ядер переходит в дочернюю особь (рис. 31). На этой стадии обе особи оказываются еще связанными друг с другом мостиком цитоплазмы и обе раковинки (старая и вновь образовавшаяся) направлены одна к другой устьями. Вскоре после этого цитоплазматический мостик между особями утончается и перешнуровывается и обе корненожки переходят к самостоятельному существованию. По существу этот процесс мало чем отличается от деления амеб, он несколько усложнен лишь процессом образования новой раковинки.

Рис. 31. Бесполое размножение путем деления раковинной корненожки Euglypha alveolata: А - корненожка перед делением; Б-образование цитоплазматической почки, на поверхности которой располагаются скелетные пластинки; В - деление ядра, скелетные пластинки образуют новую раковину; Г-конец деления, одно из ядер переместилось в дочернюю особь. 1 - ядро; 2 - псевдоподии

Таблица 2. Различные радиолярии, зарисованные с живых объектов с их естественной окраской: 1 - Acanthodesmia prismatium (отряд Nasselaria). Тонкие радиально расходящиеся псевдоподии и желтые сферические симбионты. Кремневый скелет в форме трех колец с короткими отростками; 2 - Euchitonia virchovi (отряд Spumellaria). Многочисленные тонкие псевдоподии, сетчатый трехлопастный кремневый скелет; цитоплазма окрашена в красный цвет благодаря пигменту; 3 - Auloceras arborescens (отряд Pheodaria). Коричневая центральная капсула, зеленый феодий; кремневый скелет в форме радиально ветвящихся на концах игл и поверхностно расположенных тонких иголочек (спикул); 4 - Diplocercus fuscus (отряд Acantharia). Нитевидные псевдоподии, скелет, состоящий из сернокислого стронция из радиальных неравномерно развитых игл; зеленые симбионты зоохлореллы; 5 - Arachnocorys circumtexta (отряд Nasselaria). Нитевидные псевдоподии, кремневый скелет в форме шлема с расходящимися иглами; красная центральная капсула, желтые симбионты; 6 - Tuscarilla nationalis (отряд Pheodaria). Нитевидные псевдоподии, кремневый скелет в форме конуса с отходящими от него иглами, две центральные капсулы, темно-зеленый феодий; 7 - Lithoptera miilleri (отряд Acantharia). Скелет из сернокислого стронция в форме радиальных неравномерно развитых игл с сетчатыми выростами на концах, центральная капсула крестообразной формы с зелеными симбионтами зоохлореллами; 8 -Acanthometra tetracopa (отряд Acantharia). Немногочисленные радиальные псевдоподии; скелет состоит из сернокислого стронция, слагается из 20 радиально расположенных одинаково развитых игл; цитоплазма прикрепляется к иглам при помощи сократимых волоконец (миофрисков). В центре ярко окрашенная центральная капсула с зернами пигмента и зоохлореллами
Как уже говорилось выше, раковинные корненожки - обитатели пресных вод. Они входят в состав донного населения, причем большая часть видов приурочена к прибрежной зоне. Преимущественно это обитатели мелких стоячих водоемов - прудов, канав, богатых органическими веществами.
Довольно богатая фауна корненожек (несколько десятков видов) встречается в сфагновых болотах, в самом сфагновом мху. Этот мох очень гигроскопичен и всегда впитывает большое количество воды. В прослойках воды, между стебельками и листочками мха, живут многочисленные раковинные корненожки. Здесь же встречаются и некоторые виды инфузорий.
Таким путем создается чрезвычайно характерный биоценоз обитателей сфагновых мхов.
Отряд фораминиферы (Foraminifera)
Самым обширным отрядом среди корненожек являются обитатели моря - фораминиферы (Foraminifera). В составе современной морской фауны известно свыше 1000 видов фораминифер. Небольшое число видов, представляющих, вероятно, остаток морской фауны, обитает в подпочвенных соленых водах и солоноватых колодцах Средней Азии.
Подобно раковинным корненожкам, все фораминиферы имеют раковину. Строение скелета достигает здесь большой сложности и огромного разнообразия.
В океанах и морях фораминиферы распространены повсеместно. Их находят во всех широтах и на всех глубинах, начиная от прибрежной литоральной зоны и кончая самыми глубокими абиссальными впадинами. Все же наибольшее разнообразие видов фораминифер встречается на глубинах до 200-300 м. Подавляющее большинство видов фораминифер является обитателями придонных слоев, входят в состав бентоса. Лишь очень немногие виды живут в толще морской воды, являются планктонными организмами.
Познакомимся с некоторыми наиболее характерными формами скелета фораминифер (рис. 32).

Рис. 32. Раковинки различных фораминифер: 1 - Saccamina sphaerica; 2 - Lagena plurigera; 3 - Hyperammina elongata; 4 - то же в разрезе; 5 - Rhabdam-mina linearis; в - то же в разрезе; 7 - Astrorhiza limicola; 8 - Ammo discus incertus, вид сбоку; 9 - то же со стороны устья; 10 - Cornuspira involvens; 11 - Rheopax nodulosus; 12 - Nodosaria hispida; 13 - Haplophragmoides canariensis, вид сбоку; 14 - то же со стороны устья; 15 - Nonion umbilicatulus; 16 - то же со стороны устья; 17 - Discorbis vesicularis; 18 - тоже, вид со стороны основания; 19 - Quinqueloculina seminulum (вид сбоку); 20 -то же со стороны устья; 21 - Spiroloculina depressa; 22- Textularia sagittula; 23- Globigerina sp
Среди огромного разнообразия строения раковин фораминифер можно различить по составу их два типа. Одни из них состоят из посторонних телу корненожки частиц - песчинок. Подобно тому как мы это видели у диффлюгии (см. рис. 30), фораминиферы, обладающие такими агглютинированными раковинами, заглатывают эти посторонние частицы, а затем выделяют их на поверхности тела, где они закрепляются в тонком наружном кожистом слое цитоплазмы. Такой тип строения раковины имеют часто встречающиеся представители родов Hyperammina, Astrorhiza (см. рис. 32, 3-7) и др. Например, в некоторых районах наших северных морей (море Лаптевых, Восточно-Сибирское море) эти крупные фораминиферы, достигающие 2-3 см длины, почти сплошным слоем покрывают дно.
Число видов фораминифер с агглютинированной раковиной относительно невелико (хотя число особей этих видов может быть огромным). Большая часть обладает известковыми раковинами, состоящими из углекислого кальция (СаСО3). Эти раковины выделяются цитоплазмой корненожек, которые обладают замечательной особенностью концентрировать в своем теле кальций, содержащийся в морской воде в небольших количествах (соли кальция в морской воде составляют немногим более 0,1%). Размеры известковых раковин разных видов фораминифер могут быть очень различны. Они варьируют в пределах от 20 мк до 5-6 см. Это примерно такое же соотношение размеров, как между слоном и тараканом. Наиболее крупных из фораминифер, раковина которых имеет 5-6 см в диаметре, уже нельзя назвать микроскопическими организмами. Наиболее крупные (роды Cornuspira и др.) живут на больших глубинах.
Среди известковых раковинок фораминифер в свою очередь можно различить две группы.
Однокамерные фораминиферы имеют одну-единственную полость внутри раковины, которая сообщается с наружной средой устьем. Форма однокамерных раковин разнообразна. У одних (например, Lagena) раковинка напоминает бутылочку с длинным горлышком, иногда снабженную ребрами (см. рис. 32, 2).
Очень часто происходит спиральное закручивание раковины, и тогда внутренняя полость ее становится длинным и тонким каналом (например, Ammodiscus, см. рис. 32, 8, 9).
Большинство известковых раковин корненожек является не однокамерными, а многокамерными. Внутренняя полость раковины поделена перегородками на ряд камер, число которых может достигать нескольких десятков и сотен. Перегородки между камерами не сплошные, в них имеются отверстия, благодаря чему протоплазматическое тело корненожки не расчленено на части, а представляет собой единое целое. Стенки раковинок не у всех, но у многих фораминифер пронизаны мельчайшими порами, которые служат для выхода наружу псевдоподий. Об этом подробнее будет сказано ниже.
Число, форма и взаимное расположение камер в раковинке может быть очень различным, что и создает огромное разнообразие фораминифер (см. рис. 32). У некоторых видов камеры расположены в один прямой ряд (например, Nodosaria, см. рис. 32, 12), иногда же их расположение двурядное (Textularia, см. рис. 32, 22). Широко распространена спиральная форма раковины, когда отдельные камеры расположены по спирали, причем по мере приближения к камере, несущей устье, размеры их возрастают. Причины этого постепенного нарастания размеров камер станут понятны, когда мы рассмотрим ход их развития.
В спиральных раковинках фораминифер имеется несколько оборотов спирали. Наружные (более крупные) обороты могут быть расположены рядом с внутренними оборотами (см. рис. 32,17,18) так, что все камеры видны снаружи. Это эволютный тип раковины. У других форм наружные (более крупные) камеры целиком или частично охватывают внутренние камеры (рис. 33, 1). Это инволютный тип раковины. Особую форму строения раковины находим у фораминифер милиолид (семейство Miliolidae, рис. 32, 19). Здесь камеры сильно вытянуты параллельно продольной оси раковины и расположены в нескольких пересекающихся плоскостях. Вся раковина в целом оказывается продолговатой и по форме несколько напоминает тыквенное зерно. Устье расположено на одном из полюсов и обычно снабжено зубцом.

Рис. 33. Раковины фораминифер: 1 - Elphidium strigilata; 2 - Archiacina verworni
Большой сложностью строения отличаются раковинки, относящиеся к циклическому типу (роды Archiacina, Orbitolites и др., см. рис. 33, 2; 34). Число камер здесь очень велико, причем внутренние камеры располагаются по спирали, наружные же-концентрическими кольцами.
В чем заключается биологическое значение столь сложного строения многокамерных раковинок корненожек? Специальное исследование этого вопроса показало, что многокамерные раковинки по сравнению с однокамерными обладают гораздо большей прочностью. Основное биологическое значение раковинки - это защита мягкого протоплазматического тела корненожки. При многокамерном строении раковинки эта функция осуществляется весьма совершенно.
Как устроено мягкое протоплазматическое тело фораминифер?
Внутренняя полость раковины заполнена цитоплазмой. Внутри раковинки помещается и ядерный аппарат. В зависимости от стадии размножения (о чем будет сказано несколько ниже) ядро может быть одно или их несколько. Из раковинки через устье наружу выдаются многочисленные очень длинные и тонкие псевдоподии, ветвящиеся и анастомозирующие между собой. Эти особые свойственные фораминиферам ложные ножки называют ризоподиями. Последние образуют вокруг раковинки тончайшую сеточку, общий диаметр которой обычно значительно превосходит диаметр раковинки (рис. 34). У тех видов фораминифер, у которых имеются поры, ризоподии выдаются наружу через поры.

Рис. 34. Фораминифера Orbitolites complanatus с ризоподнями
Функция ризоподии двояка. Они являются органоидами движения и захвата пищи. К ризоподиям "прилипают" различные мелкие пищевые частицы, очень часто это бывают одноклеточные водоросли. Переваривание их может происходить двояко. Если частичка мала, она постепенно как бы "скользит" по поверхности ризоподии и через устье втягивается внутрь раковинки, где и происходит переваривание. Если пищевая частица велика и не может быть втянута внутрь раковины через узкое устье, то переваривание происходит вне раковинки. Вокруг пищи при этом собирается цитоплазма и образуется местное, иногда довольно значительное утолщение ризоподии, где и осуществляются процессы пищеварения.
Выполненные за последние годы с применением цейтраферной киносъемки исследования показали, что цитоплазма, входящая в состав ризоподии, находится в непрерывном движении. Вдоль ризоподии в центростремительном (к раковине) и центробежном (от раковины) направлениях довольно быстро текут токи цитоплазмы. По двум сторонам тонкой ризоподии цитоплазма как бы струится в противоположных направлениях. Механизм этого движения до сих пор остается не выясненным.
Размножение фораминифер происходит довольно сложно и у большинства видов связано с чередованием двух разных форм размножения и двух поколений. Одно из них бесполое, второе - половое. В настоящее время процессы эти изучены у многих видов фораминифер. Не вдаваясь в детали, рассмотрим их на каком-нибудь конкретном примере.
На рисунке 35 изображен жизненный цикл фораминиферы Elphidium crispa. Этот вид представляет собой типичную многокамерную фораминиферу со спирально закрученной раковинкой. Начнем рассмотрение цикла с многокамерной корненожки, обладающей маленькой зародышевой камерой в центре спирали (микросферическое поколение).

Рис. 35. Жизненный цикл фораминиферы Elphidium crispa: слева внизу - выход зародышей, образовавшихся в результате бесполого размножения; сверху-выход гамет и их копуляция
В цитоплазме корненожки первоначально имеется одно ядро. Бесполое размножение начинается с того, что ядро последовательно несколько раз делится, в результате чего образуется множество небольшого размера ядер (обычно несколько десятков, иногда свыше сотни). Затем вокруг каждого ядра обособляется участок цитоплазмы и все протоплазматическое тело корненожки распадается на множество (по числу ядер) одноядерных амебообразных зародышей, которые выходят через устье наружу. Сразу же вокруг амебовидного зародыша выделяется тонкая известковая раковинка, которая и явится первой (эмбриональной) камерой будущей многокамерной раковины. Таким образом, при бесполом размножении на первых стадиях своего развития корненожка является однокамерной. Однако очень скоро к этой первой камере начинают добавляться следующие. Происходит это так: из устья сразу выступает наружу некоторое количество цитоплазмы, которая тотчас же выделяет раковинку. Затем наступает пауза, в течение которой простейшее усиленно питается и масса протоплазмы его увеличивается внутри раковины. Затем вновь часть цитоплазмы выступает из устья и вокруг нее образуется очередная известковая камера. Этот процесс повторяется несколько раз: возникают все новые и новые камеры, пока раковина не достигнет характерных для данного вида размеров. Таким образом, развитие и рост раковины носит ступенчатый характер. Размеры и взаимное положение камер определяется тем, какое количество протоплазмы выступает из устья и как эта протоплазма располагается по отношению к предшествующим камерам.
Мы начали рассмотрение жизненного цикла Elphidium с раковинки, обладавшей очень маленькой эмбриональной камерой. В результате бесполого размножения получается раковинка, эмбриональная камера которой значительно крупнее, чем у той особи, которая приступала к бесполому размножению. В результате бесполого размножения получаются особи макросферического поколения, которые существенно отличаются от дающего им начало микросферического поколения. В данном случае потомство оказывается не вполне похожим на родителей.
Каким же путем возникают особи микросферического поколения?
Они развиваются в результате полового размножения макросферического поколения. Происходит это следующим образом. Как и при бесполом размножении, половой процесс начинается с деления ядра. Количество образующихся при этом ядер значительно больше, чем при бесполом размножении. Вокруг каждого ядра обособляется небольшой участок цитоплазмы, и таким путем формируется огромное количество (тысячи) одноядерных клеток*. Каждая из них снабжена двумя жгутиками, благодаря движению которых клетки активно и быстро плавают. Эти клетки являются половыми клетками (гаметами). Они сливаются друг с другом попарно, причем слияние затрагивает не только цитоплазму, но и ядра. Этот процесс слияния гамет и есть половой процесс. Образующаяся в результате слияния гамет (оплодотворения) клетка носит название зиготы. Она дает начало новому микросферическому поколению фораминиферы. Вокруг зиготы сразу же по ее образовании выделяется известковая раковина - первая (эмбриональная) камера. Затем процесс развития и роста раковины, сопровождающийся увеличением числа камер, осуществляется по тому же типу, как и при бесполом размножении. Раковина получается микросферической потому, что размер зиготы, выделяющей эмбриональную камеру, во много раз меньше одноядерных амебовидных зародышей, образующихся при бесполом размножении. В дальнейшем микросферическое поколение приступит к бесполому размножению и вновь даст начало макросферическим формам.
* (У некоторых видов фораминифер не все ядра участвуют в формировании гамет. Часть их остается в виде вегетативных ядер, не участвующих в процессах размножения. Этим вегетативные ядра напоминают макронуклеусы инфузорий.)
На примере жизненного цикла фораминифер мы встречаемся с интересным биологическим явлением закономерного чередования двух форм размножения - бесполого и полового, сопровождающегося чередованием двух поколений - микросферического (развивается из зиготы в результате оплодотворения) и макросферического (развивается из одноядерных амебоидных зародышей в результате бесполого размножения).
Отметим еще одну интересную особенность полового процесса фораминифер. Известно, что у большинства животных организмов половые клетки (гаметы) бывают двух категорий. С одной стороны, это крупные, богатые протоплазмой и запасными питательными веществами неподвижные яйцевые (женские) клетки, а с другой - мелкие подвижные сперматозоиды (мужские половые клетки). Подвижность сперматозоидов обычно связана с наличием у них активно двигающегося нитевидного хвостового отдела. У фораминифер, как мы видели, морфологических (структурных) различий между половыми клетками нет. По строению своему все они одинаковы и благодаря наличию жгутиков обладают подвижностью. Здесь нет еще структурных различий, которые позволили бы различать мужские и женские гаметы. Такая форма полового процесса является исходной, примитивной.
Как уже было сказано, огромное большинство современных видов фораминифер являются придонными (бентическими) организмами, встречающимися в морях всех широт от прибрежной зоны вплоть до самых больших глубин мирового океана. Изучение распределения корненожек в океане показало, что оно зависит от ряда факторов внешней среды - от температуры, глубины, солености. Для каждой из зон типичны свои виды фораминифер. Видовой состав фораминифер может служить хорошим показателем условий среды обитания.
Среди фораминифер имеются немногочисленные виды, ведущие планктонный образ жизни. Они постоянно "парят" в толще водяной массы. Типичный пример планктонных фораминифер - разные виды глобигерин (Globigerina, рис. 36). Строение их раковинок резко отличается от строения раковинок донных корненожек. Раковинки глобигерин более тонкостенные, а главное, несут многочисленные расходящиеся во все стороны придатки - тончайшие длинные иглы. Это одно из приспособлений к жизни в планктоне. Благодаря наличию игл поверхность тела, а именно отношение поверхности к массе - величина, называемая удельной поверхностью, возрастает. Это увеличивает трение при погружении в воду и способствует "парению" в воде.

Рис. 36. Планктонная фораминифера Globigerina bulloides
Широко распространенные в современных морях и океанах фораминиферы были богато представлены и в прежние геологические периоды начиная с самых древних кембрийских отложений. Известковые раковинки после размножения или смерти корненожки опускаются на дно водоема, где входят в состав отлагающегося на дне ила. Процесс этот совершается десятки и сотни миллионов лет; в результате на дне океана образуются мощные отложения, в состав которых входит несметное количество раковинок корненожек. При горообразовательных процессах, которые совершались и совершаются в земной коре, как известно, некоторые области дна океана поднимаются и становятся сушей, суша опускается и становится дном океана. Большая часть современной суши в различные геологические периоды была дном океана. Это относится в полной мере и к территории Советского Союза (за исключением немногих северных районов нашей страны: Кольский полуостров, большая часть Карелии и некоторые другие). Морские донные отложения на суше превращаются в горные осадочные породы. Во всех морских осадочных породах присутствуют раковинки корненожек. Некоторые же отложения, как например меловые, в основной своей массе состоят из раковин корненожек. Столь широкое распространение фораминифер в морских осадочных породах имеет большое значение для геологических работ, и в частности для геологической разведки. Фораминиферы, как и все организмы, не оставались неизменными. В течение геологической истории нашей планеты происходила эволюция органического мира. Изменялись и фораминиферы. Для разных геологических периодов истории Земли характерны свои виды, роды и семейства фораминифер. Известно, что по остаткам организмов в горных породах (окаменелостям, отпечаткам и т. п.) можно определить геологический возраст этих пород. Для этой цели могут быть использованы и фораминиферы. Как ископаемые они благодаря своим микроскопическим размерам представляют очень большие преимущества, так как могут быть обнаружены в очень небольших количествах горной породы. При геологической разведке полезных ископаемых (в особенности при разведке нефти) широко используется метод бурения. При этом получается колонка породы небольшого диаметра, охватывающая все слои, через которые прошел бур. Если эти слои представляют собой морские осадочные породы, то в них при микроскопическом анализе всегда обнаруживаются фораминиферы. Ввиду большой практической важности вопрос о приуроченности определенных видов фораминифер к тем или иным осадочным породам известкового возраста разработан с большой степенью точности.
Подкласс лучевики, или радиолярии (Radiolaria)
Еще более обширную по числу видов группу морских саркодовых, чем фораминиферы, образуют лучевики, или радиолярии (Radiolaria).
Это отдельный подкласс в классе саркодовых, насчитывающий не менее 7- 8 тыс. видов. Кроме современных видов, радиолярии богато представлены и в ископаемом состоянии. Это обусловлено тем, что у большинства их, так же как и у фораминифер, имеется минеральный скелет.