Видовой состав фитоценозов
Конституционная структура фитоценозов
Понятие конституционной структуры фитоценоза, предложенное Т.A. Работновым (1965), отражает его состав в широком смысле, включая видовой, популяционный, эколого-биологический состав, состав фитоценотипов и ценогенетических групп и т.д. Ниже рассматриваются вкратце основные черты конституционной структуры фитоценозов.
Видовой состав фитоценозов
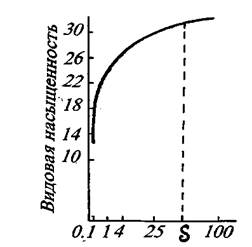
Каждый фитоценоз характеризуется особым, свойственным ему видовым составом. Сложность или простота его определяется показателем видовой (флористической) насыщенности, под которой понимают число видов, приходящееся на единицу площади фитоценоза.[1] Зависимость величины данного показателя от площади учета определяется кривой регрессии, которая вначале идет резко вверх, а затем становится более пологой (рис. 12). Характер такой кривой свидетельствует о том, что для выявления видового состава фитоценоза площадь учета не должна быть меньше определенного порогового значения (S), которое очень сильно зависит от размеров образующих ценоз растений. Поэтому при установлении видового состава сообществ разных типов используются пробные площади (площади учета) неодинаковых размеров, например лесные фитоценозы описываются обычно на пробной площади 0,25 га, травяные - 100 кв.м, моховые и лишайниковые - 1 кв. м.
По значению показателя видовой насыщенности фитоценозы можно разделить на три группы: a) флористически простые, состоящие из небольшого числа видов (до одного - двух десятков), б) флористически сложные, включающие многие десятки видов, в) фитоценозы, занимающие по видовой насыщенности промежуточное положение. Однако при определении и видовой насыщенности обычно учитываются лишь высшие растения и макрофитные лишайники. Ecли учесть, что в состав фитоценозов входят также виды водорослей, грибов, бактерий, численность которых обычно в несколько раз превышает число высших растений, то нужно признать, что в природе, по-видимому, не существует флористически простых фитоценозов, о чем свидетельствует приводимая ниже таблица 3.
|
На видовое разнообразие фитоценозов влияет ряд факторов. Определенную роль в этом плане играют общие физико-географические и исторические условия, от которых зависит видовое богатство флоры каждого конкретного района. А чем богаче флора района, тем больше будет видов-претендентов, способных поселиться в каждом конкретном фитоценозе. Так, например, видовая насыщенность фитоценозов влажных тропических лесов, формирующихся в условиях исключительно богатой тропической флоры, оценивается сотнями видов высших растений, а видовая насыщенность сибирских таежных лесов, формирующихся на фоне бедной бореальной флоры, варьирует, как правило, в пределах 15-30 видов.
Таблица 3
Полный видовой состав фитоценозов опустыненных степей Казахстана (Работнов, 1978)
| Номера фитоценозов | |||||||||
| Группы растений | Число | % | Число | % | Число | % | Число | % | |
| Цветковые | 12,5 | 10,5 | 10,5 | 12,0 | |||||
| Мхи | 0,8 | 0,5 | |||||||
| Лишайники | 4,6 | 6,0 | 9,0 | 7,0 | |||||
| Водоросли | 9,9 | 17,0 | 11,5 | 4,5 | |||||
| Микроскопические грибы | 32,3 | 34,5 | 34,0 | 39,0 | |||||
| Бактерии и актиномицеты | 39,9 | 32,0 | 34,5 | 38,0 | |||||
| Всего | |||||||||
Флористическое разнообразие фитоценозов зависит также от условий местообитаний: чем они благоприятнее, тем сложнее видовой состав, и, наоборот, в неблагоприятных местообитаниях формируются флористически простые фитоценозы. Например, в ненарушенных фитоценозах луговых степей европейской части России, связанных с умеренно увлажненными плодородными черноземными почвами, насчитывалось (Алехин, 1935) на 100 кв. м . до 120 и более видов высших растений, тогда как в этих же районах по берегам водоемов с неблагоприятными сильно переувлажненными почвами или на солончаках формируются фитоценозы из 5-10 видов высших растений.
Таким образом, неблагоприятные факторы экотопа исключают возможность произрастания в нем многих видов и создают так называемую экотопическую замкнутость фитоценоза, обусловливающую в данном случае простоту его состава. Поэтому М.В. Марков (1962) вполне справедливо рассматривал видовую насыщенность фитоценоза как показатель экологической емкости местообитания.
Кроме экотопических факторов на видовое разнообразие фитоценозов оказывают влияние также ценотические условия, что проявляется, во-первых, в конкурентном исключении одних видов растений другими и, во-вторых, в образовании в сообществах специфической внутренней среды, препятствующей внедрению в них видов, приспособленных к данной среде. Так, например, на месте уничтоженных темнохвойных лесов таежной зоны Западной Сибирской равнины формируются производные березовые леса, которых со временем береза вытесняется темнохвойными древесными породами. Вместе с ней исчезают из нижних ярусов и в дальнейшем отсутствуют некоторые ее спутники, не выносящие создаваемого в темнохвойных лесах значительного затенения обильного хвойного опада, повышенной кислотности почвы. Данное явление, названное А.К. Куркиным (1976) ценотической замкнутостью фитоценозов, широко распространено в пpироде. При этом экотопический и ценотический аспекты замкнутости взаимосвязаны друг с другом и совместно определяют общую экологическую замкнутость фитоценоза и ограничивают емкость его местообитания.
На видовое разнообразие фитоценозов могут оказывать влияние также животные и человек. Так, продолжительный интенсивный выпас домашних животных приводит к значительному упрощению видового состава исходно флористически богатых луговых и степных фитоценозов. Человек иногда сознательно уничтожает в фитоценозах нежелательные для него растения и тем самым упрощает их видовой состав. Например, так поступают при очистке от сорняков агрофитоценозов. В других случаях, напротив, человек вводит в состав естественных фитоценозов новые полезные для него растения, усложняя видовой состав. Так поступают при улучшении состава естественных пастбищ путем подсева ценных кормовых трав. Чаще же всего человек сознательно или бессознательно влияет на местообитание, изменяя его и тем самым вызывает изменение видового состава фитоценозов: удобряя луга, осушая болота, орошая естественные фитоценозы степей и пустынь.
Кроме того, видовое разнообразие фитоценоза может зависеть от режима поступления зачатков растений на его территорию извне, что в первую очередь определяется ландшафтным положением фитоценоза. Чем разнообразнее и в большем количестве поступают в фитоценоз зачатки растений со стороны, тем выше вероятность того, что данный фитоценоз будет богат флористически. И хотя эта закономерность не всегда реализуется в природе, но тенденция ее проявления сохраняется постоянно. Еще в начале двадцатых годов Л.Г. Раменский (1971) обратил внимание на то, что в естественных фитоценозах далеко не всегда имеются жизнеспособные зачатки всех тех видов растений, которые могли бы в них произрастать. Учитывая данное обстоятельство, он ввел понятия флористической полночленности и флористической неполночленности фитоценозов и предложил различать две аналогичные категории фитоценозов.
Флористически полночленные фитоценозы включают все виды, способные в них произрастать. Позднее было установлено (Работнов, 1978), что абсолютно полночленных фитоценозов, по-видимому, не существует в природе, так как в любом случае найдутся виды из других флористических областей, способные произрастать в том или ином конкретном фитоценозе. Об этом свидетельствуют факты широкого распространения и внедрения в естественные фитоценозы видов, случайно занесенных человеком из других регионов. Поэтому можно предположить лишь существование туземно-полночленных фитоценозов, которые включают все виды местной флоры, способные в них произрастать. Такие фитоценозы следует искать среди устойчивых "эколого-фитоценотические замкнутых" (Куркин, 1976) сообществ.
Явно неполночленные фитоценозы не содержат всех видов местной флоры, способных в них произрастать. При поступлении зачатков естественным путем или с помощью человека эти виды приживаются в данных фитоценозах. По-видимому, большинство природных фитоценозов являются флористически неполночленными (Василевич, 1983), о чем, в частности, свидетельствуют факты успешного искусственного введения полезных растений во многие естественные фитоценозы.
Вообще же вопрос о полночленности или неполночленности фитоценозов решается непросто, так как основывается на длительных наблюдениях за судьбой подсеянных или подсаженных новых видов, чтобы убедиться в результатах их приживания. Вместе с тем, данный вопрос имеет важное практическое значение, так как с наличием флористический неполночленности фитоценозов связана возможность сознательного введения в естественные фитоценозы полезных человеку растений, успешная борьба с сорняками, повышение продуктивности природных сенокосов и пастбищ и решение ряда других задач рационального использования растительности.
Таковы основные факторы, определяющие видовое разнообразие фитоценозов. Кроме того, имеются и некоторые другие причины, влияющие на видовую насыщенность, например, резкая переменность экологических режимов в ряде местообитаний, некоторые эколого-биологические особенности видов и т.д.
Ценопопуляции
Каждый вид в фитоценозе почти всегда бывает представлен более или менее значительным числом особей, которые в совокупности образуют ценопопуляцию. Представление о ценопопуляции было разработано в сороковых - начале пятидесятых годов ТА. Работновым (1945, 1950а и др.), а термин ценопопуляции был введен в геоботанику позже В.В. Петровским (1960) и А.А. Корчагиным (1964). В дальнейшем учение о ценопопуляции интенсивно развивалось А.А. Урановым и его учениками, опубликовавшими много интересных работ, в том числе двухтомную монографию "Ценопопуляции растений" (1975,1989).
К настоящему времени сформировалось представление о ценопопуляции, как одном из основных элементов состава фитоценоза, а сам фитоценоз нередко определяется как система ценопопуляций, связанных друг с другом и со средой. Каждая ценопопуляция занимает свою экологическую нишу. При совместном произрастании видов в фитоценозе ниши ценопопуляций частично перекрываются, но их центры всегда дифференцированы (Миркин, Розенберг, 1978). Ценопопуляции представляют собой гетерогенное образование. Она состоит из особей, различающихся по возрасту, размерам, жизненному состоянию, реакции на внешние воздействия и т.д. Считается, что внутренняя дифференциация ценопопуляции является фактором устойчивости. В связи с непрерывностью растительного покрова и слабой дискретностью естественных фитоценозов ценопуляции одного вида из соседних фитоценозов оказываются связанными постепенными, континуумообразными переходами и нечетко отграничиваются друг от друга (Миркин, Розенберг, 1983).
Каждая ценопопуляция занимает в фитоценозе определенное положение, т.е. играет определенную роль, которая зависит, прежде всего, от следующих признаков ценопопуляции: количественного участия, возрастного состава, фенологического состояния, жизненного состояния, эколого-биологических особенностей.