Характеристика умовних і безумовних рефлексів. Методи вивчення вищої нервової діяльності
Планування заходів з охорони праці
Система планів з охорони праці окремого підприємства може включати:
-Перспективне планування (на період, більший одного року);
-Поточне планування (на рік);
-Оперативне планування (детальні плани, спрямовані на вирішення конкретних питань працеохоронної діяльності на підприємстві в короткостроковому, до одного року, періоді).
Метою планування є визначення необхідних вкладень у заходи з охорони праці для ефективного впливу на стан охорони праці.
Планування в охороні праці може включати:
-Визначення цілей працеохоронної діяльності на підприємстві та засобів їх досягнення;
-Вибір методів і базових показників, за допомогою яких може здійснюватися оцінка необхідних вкладень в охорону праці;
-Розрахунок суми вкладень у заходи з охорони праці та раціональний розподіл цієї суми за напрямками працеохоронної діяльності;
-Забезпечення організації контролю виконання плану (при необхідності здійснення коригування запланованих показників);
-Здійснення постійного контролю умов і безопасностітруда на підприємстві та оперативне реагування на відхилення від нормативних вимог.
Визначення цілей планування працеохоронної діяльності на підприємстві здійснюється з урахуванням наступних пріоритетів:
-Розробка стратеги щодо безпеки праці і виробничого середовища;
-Забезпечення повноцінної організаційної структури в охороні праці;
-Вибір методів і критеріїв оцінки виконання завдань охорони праці та забезпечення функціонування систем контролю.
У процесі розробки стратегії працеохоронної діяльності виділяються чотири етапи:
-Комплексний економічний аналіз (оцінка) стану умов і охорони праці на підприємстві;
-Визначення взаємодії основних зовнішніх і внутрішніх факторів, які впливають на працеохоронної витрати підприємства;
-Розробка стратегічних альтернатив нейтралізації основних негативних чинників (забезпечення дотримання нормативних вимог охорони праці);
-Вибір стратегії, яка відповідає вимогам ефективності виробництва.
Комплексний аналіз стану умов і охорони праці передбачає:
-Оцінку того, наскільки успішною є на сьогодні стратегія працеохоронної діяльності на підприємстві;
-Визначення сильних і слабких складових сьогоднішньої стратегії, зовнішніх позитивних і негативних факторів, які впливають на неї;
-Порівняльний аналіз працеохоронної витрат і визначення майбутніх стратегічних проблем з охорони праці на підприємстві.
16.Покритонасінні або квітковірослини (Angiospermae або Magnoliophyta) — таксон вищих рослин, які формують квітки. Ця найважливіша група наземних рослин налічує понад 165 порядків, 540 родин, більше 13 000 родів і, ймовірно, не менше 250 000 видів. По числу видів квіткові рослини значно перевершують решту груп вищих рослин, узятих разом. Вони складають одну з груп насінних рослин. Найважливіша особливість квіткових рослин — наявність спеціалізованого органу, квітки, що бере на себе функції розмноження і залучення агентів запилення. Квіткові рослини формують свої насінні зачатки в порожнині зав'язі, яка утворена зрощенням колись відкритого плодолистка. Стінки зав'язі після запліднення розростаються і видозмінюються, даючи структуру під назвою плід. У решті груп насінних рослин насіннєвий зачаток не прихований від запилення, а насіння не поміщене в дійсний плід, але іноді насіння можуть покривати м'ясисті структури, наприклад, у представників роду тис.
Біологічна класифікація
Домен: Ядерні (Eukaryota)
Царство: Зелені рослини (Viridiplantae)
Відділ: Streptophyta
Streptophytina
- Ембріофіти (Embryophyta)
- Судинні (Tracheophyta)
Euphyllophyta
- Насінні (Spermatophyta)
Надклас: Покритонасінні (Magnoliophyta)
Історія поняття
Ботанічний термін «Angiospermae» (дослівно «покритонасінні») запропонував Пауль Герман (Paul Hermann) в 1690 році, термін був складений з грецьких слів αγγειον (вмістище) і σπερμα (насіння). Так Герман називав один з найголовніших відділів рослинного царства, якій включав рослини, що мають заключене в капсули насіння. Gymnospermae по Герману були квітковими рослинами, чий єдиний цілісний плід вважався насінням з відсутніми покривами. Сам термін і його антонім були підхоплені Карлом Ліннеєм, який використовував їх в схожому, але обмеженішому сенсі — для назв рядів свого класу Двусильних (Didynamia). У своєму сучасному значенні ці терміни почали використовуватися після того, як Роберт Броун в 1827 році встановив існування істинно голих насіннєвих зачатків у Cycadeae і Coniferae, привласнивши їм назву Gymnospermae. З того часу термін Angiospermae став використовуватися різними авторами, іноді з різними варіаціями, для позначення однієї з підгруп в межах дводольних рослин (розділення «явношлюбних» рослин на безсім'ядольні, одно(сім'я)дольні і дво(сім'я)дольні набуло широкого поширення дещо раніше).
Проте після того, як Вільгельм Хофмейстер описав процеси, що відбуваються в зародковому мішку квіткових рослин (1851), і зіставив їх із заплідненням таємношлюбних, стало ясно, що Gymnospermae є групою абсолютно відмінною від Angiospermae. В результаті, поняття «покритонасінні» поступово стали розглядати як синонім поняття «квіткові», і, відповідно, дводольні (Magnoliopsida, або Dicotyledones) і однодольні (Liliopsida, або Monocotyledones) — як підгрупи у складі Angiospermae. У цьому значенні поняття «покритонасінні» (Angiospermae) використовується і до цього дня.
Походження
Один з найважливіших напрямків розвитку рослинного царства — пристосування до мінливостей навколишнього середовища. Квіткові рослини є яскравим прикладом цієї лінії і домінують на земній поверхні в дану епоху. Від полюсів до екватора немає такої ділянки, де можливе рослинне життя, але не знайдено покритонасінних. Вони удосталь зустрічаються в долинах річок і чистих озерах, в меншої кількості — в солоних озерах і морях. Проте такі водні покритонасінні не є примітивними формами, а виникли шляхом пристосування наземного предка до водного середовища. До щонайширшої географічної різноманітності додається різноманітність форм і способів зростання. Банальна ряска, що покриває поверхню ставка, є крихітним зеленим пагоном з простим корінцем, вертикально зануреним у воду, і з дуже нечіткими листами і частинами стебла. Могутнє лісове дерево сторіччями розвивало свою складну систему стовбурів і гілок, покритих незліченними гілочками і листям, а під землею відповідну площу займає могутня, добре розвинена коренева система. Між цими двома крайнощами — нескінченні градації: водні і земні трави, повзучі, прямостоячі, кущі і дерева, набагато більша різноманітність, ніж серед решти груп насінних рослин, загалом відомих як голонасінні (Gymnospermae).
Перші залишки покритонасінних датуються юрським періодом і мають вік біля 140 мільйонів років. Базуючись на сучасних даних, можна припустити, що предки покритонасінних і гнетових розділилися протягом тріасового періоду (220–202 мільйонів років тому)[1]. Залишки рослин з ознаками покритонасінних з'явилися протягом юрського і раннього крейдового періоду (135-65 мільйонів років тому), але це були досить-таки нечисленні і примітивні форми. Сліди широкого розвитку і розповсюдження покритонасінних з'явилися в палеонтологічному літописі в середині крейдового періоду (близько 100 мільйонів років тому). Але вже наприкінці цього періоду покритонасінні стали домінуючою формою рослинного життя, і в багатьох викопних рослинах пізнаються представники сучасних родин (наприклад, бук, дуб, клен і магнолія).
Структура квітки
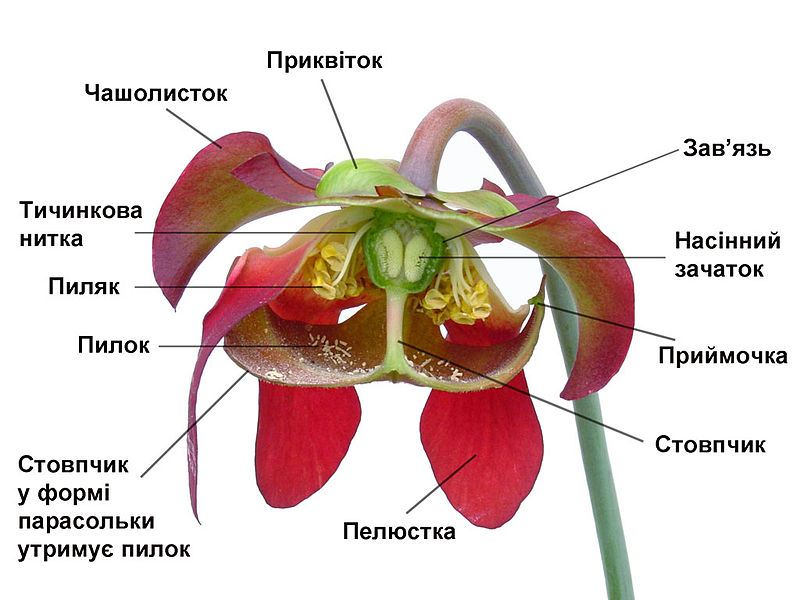
Характерною ознакою покритонасінних є квітка, яка демонструє чудову різноманітність форм і забезпечує надійні зовнішні ознаки для розрізнення видів покритонасінних. Функція квітки - забезпечення запліднення яйцеклітини і розвитку плоду, який містить насіння. Іноді, як у фіалки, квітка виникає окремо в піхві звичайного листка. Проте, звичайно, частина рослини, що несе квітку, знаходитися на кінці паростка, видається над вегетативною частиною (частиною, яка несе листя) і формує детально розроблену систему гілок, відому як суцвіття.
Відтворні (репродуктивні) клітини покритонасінних можуть бути двох видів. Одні - мікроспори, або зерна пилку, є «чоловічими» клітинами і формуються в тичинках (мікроспорофілах). Інші - мегаспори, «жіночі» клітини, в яких розвивається яйцеклітина, містяться в насінному зачатку, який знаходиться в плодолистку (мегаспорофілі). Квітка може складатися тільки з цих частин, як у верби, де кожна квітка містить тільки декілька тичинок або два плодолистки. Однак звичайно у квітці присутні стерильні структури, пристосовані як для захисту, так і для приваблення комах, необхідних для запилення. Зовнішня структура називається чашечкою і поділяється на чашолистки, які звичайно зелені та нагадують листя. Головна функція цих структур – захист квітки, особливо на стадії бруньки. Внутрішня структура називається віночком і складається з пелюсток, які загалом мають яскраві кольори і делікатнішу структуру. Її функція полягає в привабленні птахів та комах – посередників, за допомогою яких відбувається запилення. Механізми цього приваблення звичайно включають також привабливий запах та нектар, який виділяється у квітці. Ці характеристики, які існують для приваблення тварин-запилювачів, роблять квіти такими популярними серед людей. У деяких рослин, наприклад магнолії, чашолистки і пелюстки не відрізняються одне від одного, у такому разі всі вони називаються чашолистками.
Тоді як більшість квіток повноцінні, або гермафродити, тобто містять як чоловічі, так і жіночі частини в одній структурі, квіткові рослини розвинули численні морфологічні і фізіологічні механізми, щоб зменшити можливість або запобігти самозаплідненню. Гетероморфні квіти мають короткі плодолистки і довгі тичинки або навпаки, щоб перешкодити тваринам-запилювачам занести пилок в маточку (сприйнятливу частину плодолистка). Гомоморфні квіти використовують біохімічний (фізіологічний) механізм, який називається само-несумісністю, щоб відрізнити свій пилок від чужого. У деякий видів чоловічі і жіночі частини морфологічно відокремлені, розвиваючись на різних квітках.
Життєвий цикл покритонасінної рослини

Подвійне запліднення – процес у квіткових рослин протягом відтворення, при якому дві клітини сперми запліднюють дві клітини в яєчнику. Зерно пилку приклеюється до плодолистка і вирощує пилкову трубку, яка проникає у яйце через крихітну пору, мікропілу. Два спермії випускаються в яєчник через цю трубку. Одна з двох клітин сперми запліднює яйцеклітину, формуючи диплоїдну зиготу або ембріон, також відому як насінний зачаток. Друга клітини сперми зливається з двома гаплоїдними полярними ядрами в центрі ембріонального мішечка. В результаті формується триплоїдна клітина (3n). Ця клітина поділяється через мітоз і формує ендосперму, багату поживними речовинами тканину насіння. Якщо насіння розвивається без запліднення, цей процес називається апомікс.
Плід і насіння
Протягом того, як розвиток ембріона і ендосперми протікає в межах мішечка ембріона, його стіна збільшується і звичайно поглинає ядро (яке також збільшується), формуючи оболонку насіння. Стінка насінного зачатку також розвивається, формуючи плід, структуру, тісно пов'язану з розповсюдженням насіння. Часто запліднення впливає на інші частини квітки, які беруть участь в утворенні плоду, як квітколоже в яблуці, суницях та інших. Функція оболонки насіння подвійна - захисту ембріона і допомоги в розповсюдженні, вони також можуть безпосередньо допомагати проростанню. Якщо плід розкривається і тому насіння буде залишене незахищеним, оболонка забезпечує захист ембріона і, можливо, також розповсюдження. Якщо плід не розкривається, оболонка насіння майже не розвивається.
Квіткові рослини зараз розглядаються як невизначена таксономічна категорія рангом між відділом та класом. Оскільки ця таксономічна категорія більш високого рангу, ніж родина, є певна свобода у виборі назви. Стаття 16 Міжнародного кодексу ботанічної номенклатури (ICBN) дозволяє використовувати як і традиційні історичні назви, так і назву, утворену від роду. Офіційна уніномінальна назва цього таксона — Magnoliophyta, від назви роду Magnolia (Магнолія). Але традиційно укорінилися такі імена як Angiospermae і Anthophyta (квіткові рослини).
Завдяки постійному перегляду поглядів на спорідненість квіткових рослин, внутрішня систематика цієї групи піддавалася і піддається змінам. Дві широко використовувані, хоч і дещо застарілі, системи квіткових рослин — система Тахтаджяна і система Кронквіста. На сьогодні обидві ці класифікації не відображають філогенію таксона. Сучасна визнана класифікація створена міжнародною «Групою філогенії Покритонасінних» (англ. Angiosperm Phylogeny Group), яка опублікувала свій перший варіант таксономії у 1998 (відомий як APG-I), другий у 2003 (APG-II), і третій у 2009 (APG-III). Останні системи зберігають цю систематику, додаючи кілька незначних таксонів.
Традиційно, квіткові рослини були розбиті на дві групи, які в системі Кронквіста названі Magnoliopsida (у ранзі класу, сформованого від фамільного імені Magnoliacae) і Liliopsida (у ранзі класу, сформованого від фамільного імені Liliaceae). Інші описові імена, дозволені згідно зі статтею 16 ICBN, включають Dicotyledones або Dicotyledoneae, і Monocotyledones або Monocotyledoneae, які мають довгу історію використання. Для членів першої групи також може використовуватися термін «дводольна рослина», для другої – «однодольна», або «дікот» та «монокот» відповідно. Ці назви походять від факту, що дводольні звичайно мають у кожній насінині дві сім'ядолі (ембріональні листки), тоді як однодольні звичайно мають тільки одну. З діагностичної точки зору число сім'ядоль не є ні особливо зручною, ні надійною ознакою.
Недавні дослідження, проведені групою APG, показали, що однодольні — «хороша» (тобто монофілетична) група, цей таксон отримав назву «монокоти» (monocots), а дводольні – ні (вони створюють парафілетичну групу). Проте, в межах дводольних існує «хороша» група, яка включає більшість дводольних. Цей таксон називається «еудікоти» (eudicots) або «tricolpates» (від виду пилку, знайденого у представників цієї групи). Назва eudicots походить від слова «dicot» з префіксом "eu-" (від грецької 'eu'= «істина»), тобто означає «справжні дводольні», оскільки еудікоти мають ознаки, традиційні для дікотів, наприклад квіти з чотирьох або п'яти частин (чотирьох або п'яти пелюсток, чотирьох або п'яти чашолистків). Групу, яка залишається після відділення еудікотів з решти (колишніх) дікотів, іноді неофіційно називають «палеодікоти» (palaeodicots) (грецький префікс "palaeo-" означає «старий»). Оскільки представники цього залишку не створюють «хорошу» групу, цей термін існує тільки для зручності.
17.Голкошкі́рі (Echinodermata) — другий за кількістю видів (після хордових) тип вторинноротих тварин, що містить близько 6 300 сучасних видів. Усі вони — вільноживучі морські придонні тварини, що дуже чутливо реагують на опріснення. Майже всі, крім кількох батипелагічних (таких, що мешкають у товщі води на великих глибинах) голотурій, представляють собою типові бентосні форми. В батіальній та ультраабісальній зонах на частку голкошкірих може приходитися до 90 % загальної біомаси бентосу. Деякі морські зірки та голотурії зустрічаються лише на глибинах більше 2000 м.
Біологічна класифікація
Домен: Еукаріоти (Eukaryota)
Царство: Тварини (Animalia)
Підцарство: Справжні багатоклітинні (Eumetazoa)
- Двобічно-симетричні (Bilateralia)
- Вториннороті (Deuterostomia)
Тип: Echinodermata
Класи
Морські зірки (Asteroidea)
Офіури (Ophiuroidea)
Морські їжаки (Echinoidea)
Голотурії (Holothuroidea)
Морські лілії (Crinoidea)
† Бластоїдеї (Blastoidea)
Походження
Голкошкірі відомі з раннього кембрію. Особливо багато вони були представлені в пізньому палеозої. Тоді ж з'явилася і більшість груп, що дійшли до нас у викопному стані, і є більш чи менш далекими родичами сучасних форм. Завдяки хорошому збереженню їх вапнякових скелетів відомо біля 10 000 викопних видів голкошкірих. З близько 20 класів до нашого часу дожило лише шість: морські лілії Crinoidea (біля 620 видів), морські зірки Asteroidea (близько 1500 видів), офіури або змієхвостки Ophiuroidea (близько 2000 видів), голотурії або морські огірки Holothuroidea (приблизно 1200 видів) та морські їжаки Echinoidea (біля 950 видів). Раніше виділяли в окремий клас також групу Concentricycloidea (англ.) (морські маргаритки), відкриту лише у 1986 році та відому по трьом глибоководним видам, об'єднаним в однин рід Xyloplax. За сучасними уявленнями, вона входить до складу класу морських зірок.
Зовнішня будова
Голкошкірі мають, як правило, радіальну, здебільшого п'ятипроменеву, симетрію, яка дещо порушується елементами білатеральної симетрії. Радіальна симетрія голкошкірих є вторинним явищем. На це вказує двобічна симетрія личинок голкошкірих та будова ряду органів сучасних видів, а також деякі викопні форми.
У тілі голкошкірих розрізняють оральну та аборальну сторони. Оральна — та, на якій розташований ротовий отвір.
Руки голкошкірих (у морських їжаків — сектори, на яких розташовані амбулакральні ніжки) називаються радіуси. На оральній стороні кожного радіуса знаходяться амбулакральні ніжки, за доповогою яких тварина рухається (вони дозволяють побачити радіуси у морських їжаків, у яких нема рук). Сектори, що розташовані між радіусами, називаються інтеррадіуси. Зовні радіальну симетрію порушує мадрепорова пластинка, розташована на одному з інтеррадіусів.
Ендоскелет
У підшкірному шарі Голкошкірих розвивається біомінеральний ендоскелет, який складається з вапняних платівок, що є мікропористими монокристалами вапняку. Подібна структура скелетного матеріалу (стереом) притаманна лише голкошкірим. Скелет часто утворює різноманітні зовнішні придатки: голки, шипи і педицеляріі. У багатьох морських їжаків ці придатки розвиваються особливо сильно. Голки виконують захисну функцію. Часто вони бувають рухливими. Деякі морські їжаки направляють голки в бік наближення небезпеки. Педицелляріі — це голки, видозмінені в хапальні щипчики. З їх допомогою тварина чиститься і позбавляється від паразитів. У морських їжаків скелет бере участь у формуванні особливого жувального органу — аристотелевого ліхтаря. У офіур скелет особливо розвинений в променях, де він утворює ряд масивних вапняних члеників — хребців. У голотурій скелет, як правило, редукований до окремих дрібних пластинок чи голок (спікул).
Покриви
До складу епідерми Голкошкірих входять механорецепторні клітини, що забезпечують відчуття дотику, пігментні клітини, що обумовлюють забарвлення, і залозисті клітини, що виділяють липкий секрет або навіть токсини. Однією з найоригінальніших рис будови Голкошкірих є складна диференціація целому на ряд систем: амбулакральну і перигемальну системи.
Амбулакральна система унікальна серед всього царства тварин. Це мережа каналів, заповнена рідиною, за складом близькою до морської води і сполучена з навколишнім середовищем через кам'янистий канал і мадрепорову платівку. Від радіальних амбулакральних каналів відходить безліч амбулакральних ніжок, в основі яких знаходяться ампули — м'язові бульбашки, при скороченні яких ніжка подовжується. На кінці ніжки знаходиться присосок. Амбулакральна система бере участь в диханні, пересуванні та добуванні їжі. Так, за допомогою спільної роботи багатьох амбулакральних ніжок морська зірка може розкрити черепашку двостулкового молюска.
Перигемальна система — це сукупність каналів і порожнин (синусів), що оточують кровоносну систему тварини. Кровоносна система слабко розвинена і являє собою систему порожнин в сполучній тканині (лакун), що не мають ендотеліальної вистилки. У кожному промені знаходяться два радіальних перигемальних канали, в перегородці між якими розташовується радіальна кровоносна судина. Радіальні судини впадають у оральне кровоносне кільце, що лежить в перегородці між двома кільцевими перигемальними каналами. Статевий синус оточує аборальне кровоносне кільце і статевий столон. Два кровоносних кільця пов'язані осьовим органом, оточеним лівим і правим осьовими синусами.
В одному з інтеррадіусів голкошкірих розташований осьовий комплекс органів. До його складу входять органи з різних систем:
кам'янистий канал, який з'єднує кільцевий амбулакральний канал з мадрепоровою платівкою;
осьовий орган, усередині якого розташовується мережа кровоносних судин;
лівий осьовий синус — частина целому, що з'єднує внутрішній кільцевий перигемальний канал з правим осьовим синусом;
правий осьовий синус, здатний ритмічно скорочуватися і цим сприяти руху крові в судинах, тобто виконує функції серця;
статевий синус — ділянка целому, що містить статевий тяж, який складається з незрілих статевих клітин.
Нервова система
Нервова система голкошкірих примітивна, складається з трьох окремих частин, побудованих за радіальним планом: нервове кільце і радіальні нервові тяжі. В оральній стінці тіла лежать дві нервові системи — чутлива і рухова. У аборальній стінці тіла — тільки рухова. Органи чуттів голкошкірих досить різноманітні, але примітивні за будовою. Вони дифузно розподілені по тілу у вигляді різних чутливих клітин (функції дотику, хімічного чуття, зору). Світлочутливі клітини можуть бути зібрані в очах. У морських зірок очі розташовані на кінцях променів, а в морських їжаків — навколо анального отвору.
Розмноження
Більшість голкошкірих — роздільностатеві тварини. Самки утворюють багато дрібних, бідних жовтком яєць і викидають їх у воду. Таким чином, запліднення у голкошкірих зовнішнє. Розвиток зародка відбувається у воді, у складі меропланктона. Запліднена яйцеклітина (зигота) починає дробитися і через деякий час утворює бластули. Дроблення повне, радіального типу.
Зазвичай голкошкірі вилуплюються з яйця на стадії бластули. Така бластулоподібна личинка являє собою міхур, стінка якого складається з одного епітелізованого шару джгутиконосних клітин (бластомерів), а порожнина (бластоцель) заповнена драглистою рідиною. Бластула здатна пересуватися за допомогою джгутиків. Часто на анімальному полюсі личинки джгутики довші і виконують сенсорну функцію (апікальний орган).
Через деякий час на вегетативному полюсі відбувається виселення (імміграція) деяких клітин в бластоцель. Це клітини первинної мезенхіми, що беруть участь в утворенні личинкового скелета. Після цього відбувається інвагінація вегетативної стінки в бластоцель, в результаті чого формується сліпо замкнута первинна кишка (архентерон). Одночасно з цим з вершини архентерона в бластоцель виселяються клітини вторинної мезенхіми, які здатні до амебоїдного руху і сприяють інвагінації, а згодом — відокремлення целому, утворення рота та дорослих рук. Потім відбувається відокремлення целомічної мезодерми, що входить до складу архентерона, шляхом утворення випинання стінки, яке відшнуровується у формі целомічного мішка, тобто ентероцельним способом. Надалі целом ділиться спочатку на правий і лівий, а потім — на три пари целомів: праві і ліві аксоцелі (передні), гідроцелі (середні) і соматоцелі (задні). Зазвичай ліві целоми випереджають у розвитку праві, що пов'язано з їх провідною роллю в метаморфозі. На цьому завершується процес гаструляціі, в результаті якого відокремлюються три зародкових листка: ектодерма (личинкові покриви), ентодерми (кишка) і мезодерма (первинна і вторинна мезенхіма та целом). Бластопор зміщується на черевну сторону і стає анусом. На передньому кінці, на черевній стороні утворюється інвагінація ектодерми, так звана ротова бухта (стомодеум), яке зливається з кишкою і утворює рот. Кишка розчленовується на три відділи: стравохід, розширений шлунок і тонку кишку. Тіло набуває яйцеподібної форми. Навколо рота утворюється навколоротова западина, на краю якої формується війковий шнур, на решті поверхні тіла війки зникають. За допомогою биття війок клітин війкового шнура личинка пересувається і підганяє їжу до рота. Ця спільна для більшості голкошкірих білатерально-симетрична личинка отримала назву диплеврула.
Надалі личинка набуває пристосувань до планктонного способу життя, різних у різних класів, і перетворюється на плутеус (морські їжаки і офіури), аурикулярію (голотурії) або біпінарію, а пізніше — на брахіолярію (морські зірки). У видів з великою кількістю жовтка з яйця розвивається лецитотрофна (тобто така, що харчуються жовтком) бочкоподібна личинка доліолярія, підперезана поперечними війковими кільцями. Через деякий час вільноплаваюча білатерально-симетрична личинка приступає до метаморфозу, в результаті якого трансформується в радіально-симетричну дорослу тварину. У тілі личинки утворюється зачаток майбутньої дорослої тварини (імагінальний диск). На лівій стороні личинки формується оральна сторона тварини, а на правій — аборальна. До кінця метаморфозу відбувається повна редукція личинкових органів.
Характеристика умовних і безумовних рефлексів. Методи вивчення вищої нервової діяльності
І.П. Павлов усі рефлекторні реакції поділив на дві групи: безумовні і умовні. Вони ж лежать в основі поведінки людини.
Безумовні рефлекси - природжені, відносно постійні, стереотипні реакції організму на дію адекватного подразника зовнішнього або внутрішнього середовища, які здійснюються за допомогою ЦНС, передаються спадково. Їх ознаки:
1. Проявляються при дії адекватного подразника без особливих спеціальних умов (слиновиділення, ковтання, дихання та ін.).
2. Мають готові анатомічно сформовані рефлекторні дуги.
3. В їх здійсненні основна роль належить підкірковим ядрам, стовбуру мозку, спинному мозку. Вони зберігаються і після видалення кори великого мозку. Проте представництво безумовного рефлексу є в корі великих півкуль.
4. Є видовими реакціями, характерними для усіх представників даного виду.
5. Є відносно постійними рефлекторними реакціями, стійкі, незмінні, зберігаються протягом усього життя.
Безумовні рефлекси за характером реакції-відповіді поділяють на рухові, секреторні і трофічні, а за біологічною спрямованістю:
- рефлекси, пов’язані з регуляцією процесів життєдіяльності, - ковтання, жування, смоктання, слиновиділення, дихальні, серцеві, судинні тощо;
- рефлекси, пов’язані із збереження виду, - копуляція, вигодовування та піклування про потомство;
- захисні - кашель, чхання, моргання тощо;
- орієнтувальні - виникають кожного разу при дії незнайомих подразників.
Інстинкти - складні природжені безумовно-рефлекторні реакції, які проявляються за рахунок активності підкіркових ядер. Розрізняють 4 види найпростіших інстинктів : материнський, їжездобувний, статевий, захисний. Особливістю інстинктів є те, що вони викликаються внутрішніми мотивами. У їх регуляції велике значення відіграють гормони.
Безумовні рефлекси визначають певну, чітко окреслену програму поведінки, яка забезпечує пристосування до стабільних, характерних для даного виду умов життя. У зв’язку з цим тільки за рахунок безумовних реакцій неможливо пристосуватися до постійно мінливих умов навколишнього середовища.
Умовні рефлекси - індивідуальні, набуті рефлекторні реакції, які виробляються на базі безумовних рефлексів. Їх ознаки:
1. Набуваються протягом усього життя організму.
2. Неоднакові у представників одного виду.
3. Не мають готових рефлекторних дуг.
4. Вони формуються при певних умовах.
5. В їх здійсненні основна роль належить корі великого мозку.
6. Мінливі, легко виникають і легко зникають залежно від умов, в яких знаходиться організм.
Умови утворення умовних рефлексів:
1. Одночасна дія двох подразників : індиферентного для даного виду діяльності, який в подальшому стає умовним сигналом, і безумовного подразника, який викликає певний безумовний рефлекс.
2. Дія умовного подразника завжди випереджує дію безумовного (на 1-5с.).
3. Підкріплення умовного подразника безумовним повинно бути кількаразовим.
4. Безумовний подразник повинен бути біологічно сильним, а умовний володіти помірною оптимальною силою.
5. Умовні рефлекси швидше і легше формуються при відсутності сторонніх подразників.
Умовні рефлекси можна виробляти не лише на основі безумовних, але і на основі раніше набутих умовних рефлексів, які стали достатньо міцними. Це умовні рефлекси вищого порядку.
Умовні рефлекси є:
- природні - рефлекторні реакції, які виробляються на зміни навколишнього середовища, і завжди супроводять появу безумовного. Наприклад, запах, вигляд їжі є природними сигналами самої їжі;
- штучні - умовні рефлекси, що виробляються на подразнення, які не мають до безумовно рефлекторної реакції природного відношення. Наприклад, слиновиділення на дзвоник або на час.
Метод умовних рефлексів - метод дослідження ВНД. І.П. Павлов звернув увагу на те, що діяльність вищих відділів головного мозку не тільки пов’язана з прямим впливом подразників, які мають біологічне значення для організму, а й залежить від умов, які супроводять ці подразнення. Наприклад, у собаки слиновиділення починається не лише тоді, коли їжа потрапляє в рот, а й при вигляді, запахові їжі, як тільки вона побачить людину, яка завжди їй приносить їжу. І.П. Павлов пояснив це явище, розробивши метод умовних рефлексів. За методом умовних рефлексів він проводив досліди на собаках з фістулою вивідного протоку привушної слинної залози. Тварині пропонували два подразника: їжа - подразник, який має біологічне значення і викликає слиновиділення ; другий - індиферентний для процесу живлення (світло, звук). Ці подразники поєднували в часі так, щоб дія світла (звуку) на кілька секунд випереджала приймання їжі. Після ряду повторень слина починала виділятися при спалах лампочки і відсутності їжі. Світло (індиферентний подразник) назвали умовним, оскільки він є умовою, за якої проходило приймання їжі. Подразник, який має біологічне значення (їжа) назвали безумовним, а фізіологічну реакцію слиновиділення, яка відбувається внаслідок дії умовного подразника - умовним рефлексом.
Щоб з’ясувати механізм утворення умовних рефлексів, використовують часткове виділення певних частин кори великого мозку та реєстрацію електричної активності різних мозкових структур під час дії безумовного і умовного подразників.
І.П. Павлов вважав, що при одночасній дії на два різні аналізатори в різних чутливих ділянках півкуль великого мозку виникає збудження, а з часом, між ними утворюється зв’язок. Наприклад, при спалахуванні лампочки і підкріпленні цього подразника їжею виникає збудження в кірковій частині зорового аналізатора, що знаходиться в потиличній ділянці кори і збудження харчового центра кори півкуль великого мозку - тобто в обох кіркових центрах (зоровому і харчовому), між якими утворюється нервовий зв’язок, який при багаторазовому поєднанні в часі цих подразників стає міцним.
При умовних рефлексах, як і при безумовних, має місце зворотна афференція, тобто сигнал про те, що відбулась умовнорефлекторна реакція. Вона дає можливість ЦНС оцінити поведінкові акти. Без такої оцінки неможливе тонке пристосування поведінки до постійно змінних умов середовища.
Дослідження тварин, в яких видаляли ділянки кори, показало, що в цих тварин можна виробити умовні рефлекси. Отже, умовні рефлекси формуються внаслідок взаємодії кори великого мозку і підкіркових центрів. Структура рефлекторної дуги умовного рефлексу має складний характер. Так, в утворенні складних поведінкових реакцій кора має провідне значення, а при формуванні вегетативних умовних рефлексів кора і підкіркові структури відіграють однакову роль. Доведено, що руйнування сітчастого утвору затримує утворення умовних рефлексів, а подразнення його електричним струмом прискорює їх утворення.
Біологічне значення умовних рефлексів полягає в тому, що вони є пристосувальними реакціями організму, які формуються умовами життя людини і дають можливість заздалегідь пристосуватись до нових умов. Умовні рефлекси мають попереджувальне сигнальне значення, оскільки організм починає реагувати цілеспрямовано до того, як почне діяти життєво важливий подразник. Тому умовні рефлекси забезпечують живій істоті можливість заздалегідь оцінити небезпеку або корисний подразник.
19.М’язова система – частина опорно-рухової системи. У нашому тілі близько 600 м’язів. За місцем розташування вони поділяються на м’язи голови, шиї, тулуба, кінцівок. М’язи виконують безліч функцій. Рухаючи кістки, до яких вони прикріплені, м’язи забезпечують переміщення тіла у просторі й підтримку його рівноваги, дають можливість прибирати різні пози, вимовляти різноманітні звуки, здійснювати жувальні й ковтальні рухи. М’язова маса становить приблизно 50 % загальної маси тіла.
Кістякові м’язи — найбільша група м’язів нашого тіла. Це і найбільші м’язи. Вони беруть участь у різноманітних рухах тулуба й кінцівок. Кістякові м’язи утворені поперечносмугастою м’язовою тканиною. У її волокнах є елементи (міофібрили), що забезпечують одночасне скорочення одних і розслаблення інших м’язів. М’язові волокна розташовуються паралельно одне одному й об’єднані в пучки, зв’язані прошарком зі сполучної тканини. Кінці пучків переходять у сухожилля, за допомогою яких м’язи прикріплюються до кісток кістяка.
Кістякові м’язи йдуть від однієї кістки до іншої через суглоб, що їх з’єднує, і працюють парами, але у протилежному напрямку. М’яз-згинач має парний м’яз-розгинач, привідний м’яз працює в парі з відвідним м’язом і т. д. Усі види рухів м’язи-антагоністи виконують спільно, скорочуючись по черзі. Навіть у найпростіших рухах беруть участь кілька м’язів. 
Рухами м’язів, що зроблені свідомо (згинання й розгинання спини, рух пальців під час гри на музичному інструменті тощо), керує головний мозок. У його правій півкулі є вищі рухові центри, що відповідають за роботу певної групи м’язів (тулуба, голови, пальців рук тощо). Завдяки нервовим закінченням, розташованим у м’язовій тканині, скорочення м’язів може відбуватися й без участі головного мозку. Рухами м’язів, здійснюваними рефлекторно (незалежно від свідомості), керує спинний мозок, у якому містяться виконавчі нервові клітини (нейрони). У відповідь на подразнення нервових закінчень, які містяться у м’язі, у нейроні спинного мозку виникає хвиля збудження (нервовий імпульс). Довгим відростком нейрона (аксоном) імпульс повертається до м’яза й викликає його мимовільне скорочення (здійснюється безумовний рефлекс). Шлях, який проходить імпульс від нервових закінчень м’яза до спинного мозку й назад до м’яза, називається рефлекторною дугою. Витрати енергії під час роботи м’язів великі. Ії запас м’язи накопичують у власних клітинах, витягаючи поживні речовини із крові, якою вони добре забезпечуються. Якщо м’язи працюють занадто довго (біг на довгі дистанції, гра у футбол тощо), у них накопичується молочна кислота, що викликає відчуття болю. Після відпочинку біль минає. Прискорити цей процес можна, прийнявши гарячу ванну або душ і зробивши масаж м’язів. Так вчиняють спортсмени після тренувань.
Сухожилля – це дуже міцні тяжі зі сполучної тканини. Вони складаються з окремих волокон, зібраних у пучки. На одному кінці м’яза волокна сухожилля переплітаються з м’язовими волокнами і йдуть у глиб м’яза, а на іншому – вплітаються в окістя. Довгі м’язи зазвичай закінчуються циліндричними сухожиллями, а короткі м’язи – пластинчастими. Сухожилля згиначів пальців кисті й стопи оточені оболонками (синовіальними піхвами). Під час здійснення пальцями ковзних рухів вони виділяють рідину, що зменшує тертя. На відміну від зв’язок сухожилля не розтягуються, тому від скорочення м’яза кістка переміщується. Наприклад, литка кріпиться до п’яткової кістки ахіллесовим сухожиллям, що піднімає п’яту під час ходіння або бігу.