Мышечная система и ее функции
(строение, физиология и биохимия мышечных сокращений,
общий обзор скелетной мускулатуры)
Существует два вида мускулатуры: гладкая (непроизвольная) и поперечно-полосатая (произвольная). Гладкие мышцы расположены в стенках кровеносных сосудов и некоторых внутренних органах. Они сужают или расширяют сосуды, продвигают пищу по желудочно-кишечному тракту, сокращают стенки мочевого пузыря. Поперечно-полосатые мышцы – это все скелетные мышцы, которые обеспечивают многообразные движения тела. К поперечно-полосатым мышцам относится также и сердечная мышца, автоматически обеспечивающая ритмическую работу сердца на протяжении всей жизни. Основа мышц – белки, составляющие 80−85 % мышечной ткани (исключая воду). Главное свойство мышечной ткани – сократимость, она обеспечивается благодаря сократительным мышечным белкам – актину и миозину.
Мышечная ткань устроена очень сложно. Мышца имеет волокнистую структуру, каждое волокно – это мышца в миниатюре, совокупность этих волокон и образуют мышцу в целом. Мышечное волокно, в свою очередь, состоит из миофибрилл. Каждая миофибрилла разделена на чередующиеся светлые и темные участки. Темные участки – протофибриллы − состоят из длинных цепочек молекул миозина, светлые − образованы более тонкими белковыми нитями актина. Когда мышца находится в несокращенном (расслабленном) состоянии, нити актина и миозина лишь частично продвинуты относительно друг друга, причем каждой нити миозина противостоят, окружая ее, несколько нитей актина. Более глубокое продвижение относительно друг друга обусловливает укорочение (сокращение) миофибрилл отдельных мышечных волокон и всей мышцы в целом.
К мышце подходят и от нее отходят (принцип рефлекторной дуги) многочисленные нервные волокна. Двигательные (эфферентные) нервные волокна передают импульсы от головного и спинного мозга, приводящие мышцы в рабочее состояние; чувствительные волокна передают импульсы в обратном направлении, информируя центральную нервную систему о деятельности мышц. Через симпатические нервные волокна осуществляется регуляция обменных процессов в мышцах, посредством чего их деятельность приспосабливается к изменившимся условиям работы, к различным мышечным нагрузкам. Каждую мышцу пронизывает разветвленная сеть капилляров, по которым поступают необходимые для жизнедеятельности мышц вещества и выводятся продукты обмена.
Скелетная мускулатура. Скелетные мышцы входят в структуру опорно-двигательного аппарата, крепятся к костям скелета и при сокращении приводят в движение отдельные звенья скелета, рычаги. Они участвуют в удержании положения тела и его частей в пространстве, обеспечивают движения при ходьбе, беге, жевании, глотании, дыхании и т.д., вырабатывая при этом тепло. Скелетные мышцы обладают способностью возбуждаться под влиянием нервных импульсов. Возбуждение проводится до сократительных структур (миофибрилл), которые, сокращаясь, выполняют определенный двигательный акт – движение или напряжение.
Напомним, что вся скелетная мускулатура состоит из поперечно-полосатых мышц. У человека их насчитывается около 600 и большинство из них – парные. Их масса составляет 35−40 % от общей массы тела взрослого человека. Скелетные мышцы снаружи покрыты плотной соединительно-тканной оболочкой. В каждой мышце различают активную часть (тело мышцы) и пассивную (сухожилие). Мышцы делятся на длинные, короткие и широкие.
Мышцы, действие которых направлено противоположно, называются антагонистами, однонаправленно – синергистами. Одни и те же мышцы в различных ситуациях могут выступать в том и другом качестве. У человека чаще встречаются веретенообразные и лентовидные мышцы. Веретенообразные мышцы расположены и функционируют в районе длинных костных образований конечностей, могут иметь два брюшка (двубрюшные мышцы) и несколько головок (двуглавые, трехглавые, четырехглавые мышцы). Лентовидные мышцы имеют различную ширину и обычно участвуют в корсетном образовании стенок туловища. Мышцы с перистым строением, обладая большим физиологическим поперечником, за счет большого количества коротких мышечных структур значительно сильнее тех мышц, у которых ход волокон имеет прямолинейное (продольное) расположение. Первые называются сильными мышцами, осуществляющими малоамплитудные движения, вторые – ловкими, участвующими в движениях с большой амплитудой. По функциональному назначению и направлению движений в суставах различают мышцы: сгибатели и разгибатели, приводящие и отводящие, сфинктеры (сжимающие) и расширители.
Сила мышцы определяется весом груза, который она может поднять на определенную высоту или способна удерживать при максимальном возбуждении, не изменяя своей длины. Сила мышцы зависит: от суммы сил мышечных волокон, их сократительной способности; количества мышечных волокон, их сократительной способности; количества мышечных волокон в мышце и количества функциональных единиц, одновременно возбуждающихся при развитии напряжения; исходной длины мышцы (предварительно растянутая мышца развивает большую силу); условий взаимодействия с костями скелета.
Сократительная способность мышцы характеризуется ее абсолютной силой, то есть силой, приходящейся на 1 см поперечного сечения мышечных волокон. Для расчета этого показателя силу мышцы делят на площадь ее физиологического поперечника (то есть на сумму площадей всех мышечных волокон, составляющих мышцу). Например, в среднем у человека сила (на 1 см2 поперечного сечения мышцы) икроножной мышцы – 6,24 кг; разгибателей шеи – 9,0 кг; трехглавой мышцы плеча – 16,8 кг.
Центральная нервная система регулирует силу сокращения мышцы путем изменения количества одновременно участвующих в сокращении функциональных единиц, а также частотой посылаемых к ним импульсов. Учащение импульсов ведет к возрастанию величины напряжения.
Работа мышц. В процессе мышечного сокращения потенциальная химическая энергия переходит в потенциальную механическую энергию напряжения и кинетическую энергию движения. Различают внутреннюю и внешнюю работу. Внутренняя работа связана с трением в мышечном волокне при его сокращении. Внешняя работа проявляется при перемещении собственного тела, груза, отдельных частей организма (динамическая работа) в пространстве. Она характеризуется коэффициентом полезного действия (КПД) мышечной системы, то есть отношением производимой работы к общим энергетическим затратам (для мышц человека КПД составляет 15−20 %, у физически развитых тренированных людей этот показатель несколько выше). При статических усилиях (без перемещения) можно говорить не о работе с точки зрения физики, а о работе, которую следует оценивать энергетическими физиологическими затратами организма.
Мышца как орган.В целом мышца как орган представляет собой сложное образование, которое выполняет определенные функции, состоит на 72−80 % из воды и на 16−20 % из плотного вещества. Мышечные волокна состоят из миофибрилл с клеточными ядрами, рибосомами, митохондриями, саркоплазматическим ретикулюмом, чувствительными нервными образованиями – проприорецепторами и другими функциональными элементами, обеспечивающими синтез белков, окислительное фосфорилирование и ресинтез аденозинтрифосфорной кислоты, транспортировку веществ внутри мышечной клетки в процессе функционирования мышечных волокон. Важным структурно-функциональным образованием мышцы является двигательная, или нейромоторная, единица, состоящая из одного мотонейрона и иннервируемых им мышечных волокон. Различают малые, средние и большие двигательные единицы в зависимости от количества мышечных волокон, задействованных в акте сокращения.
Система соединительнотканных прослоек и оболочек связывает мышечные волокна в единую рабочую систему, обеспечивающую с помощью сухожилий передачу возникающей при мышечном сокращении тяги на кости скелета.
Вся мышца пронизана разветвленной сетью кровеносных и веточками лимфатических сосудов. Мышца состоит из красных и белых мышечных волокон. Красные мышечные волокна обладают более густой сетью кровеносных сосудов, чем белые. Они имеют большой запас гликогена и липидов, характеризуются значительной тонической активностью, способностью к длительному напряжению и выполнению продолжительной динамической работы. Каждое красное волокно имеет больше, чем белое, митохондрий – генераторов и поставщиков энергии, окруженных 3−5 капиллярами. Это создает условия для более интенсивного кровоснабжения красных волокон и высокого уровня обменных процессов.
Белые мышечные волокна имеют миофибриллы, которые толще и сильнее миофибрилл красных волокон, они быстро сокращаются, но не способны к длительному напряжению. Митохондрии белого вещества имеют только один капилляр. В большинстве мышц красные и белые волокна содержатся в разных пропорциях. Различают также мышечные волокна: тонические, способные к локальному возбуждению без его распространения; фазные, способные реагировать на распространяющуюся волну возбуждения как сокращением, так и расслаблением; переходные, сочетающие оба свойства.
Мышечный насос– физиологическое понятие, связанное с мышечной функцией и ее влиянием на собственное кровоснабжение. Принципиальное его действие проявляется следующим образом: во время сокращения скелетных мышц приток артериальной крови к ним замедляется и ускоряется ее отток по венам; в период расслабления венозный отток уменьшается, а артериальный приток достигает своего максимума. Обмен веществ между кровью и тканевой жидкостью происходит через стенку капилляра.
Функции мышц регулируются различными отделами центральной нервной системы (ЦНС), которые во многом определяют характер их разносторонней активности (фазы движения, тонического напряжения и др.). Рецепторы двигательного аппарата дают начало афферентным волокнам двигательного анализатора, которые составляют 30−50 % волокон смешанных (афферентно-эфферентных) нервов, направлящихся в спинной мозг. Сокращение мышц вызывает импульсы, которые являются источником мышечного чувства – кинестезии.
Передача возбуждения с нервного волокна на мышечное осуществляется через нервно-мышечный синапс, который состоит из двух разделенных щелью мембран – пресинаптической (нервного происхождения) и постсинаптической (мышечного происхождения). При воздействии нервного импульса выделяются кванты ацетилхолина, которые приводят к возникновению электрического потенциала, способного возбудить мышечное волокно. Скорость проведения нервного импульса через синапс в тысячи раз меньше, чем в нервном волокне. Он проводит возбуждение только в направлении к мышце. В норме через нервно-мышечный синапс млекопитающих может пройти до 150 импульсов в одну секунду. При утомлении или патологии подвижность нервно-мышечных окончаний снижается, а характер импульсов может изменяться.
Сокращение и напряжение мышцы осуществляется за счет энергии, освобождающейся при химических превращениях, которые происходят при поступлении в мышцу нервного импульса или нанесении на нее непосредственного раздражения. Химические превращения в мышце протекают как при наличии кислорода (в аэробных условиях), так и при его отсутствии (в анаэробных условиях).
Расщепление и ресинтез аденозинтрифосфорной кислоты (АТФ).Первичным источником энергии для сокращения мышцы служит расщепление АТФ (она находится в клеточной мембране, ретикулюме и миозиновых нитях) на аденозиндифосфорную кислоту (АДФ) и фосфорные кислоты. При этом из каждой грамм-молекулы АТФ освобождается 10 000 калорий:
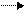 АТФ АДФ + Н3РО4 + 10 000 (кал.).
АТФ АДФ + Н3РО4 + 10 000 (кал.).
АДФ в ходе дальнейших превращений дефосфолирируется до адениловой кислоты. Распад АТФ стимулирует белковый фермент актомиозин (аденозинтрифосфотаза). В покое он не активен, активизируется при возбуждении мышечного волокна. Активность актомиозина увеличивается под воздействием ионов Са++, которые в состоянии покоя располагаются в саркоплазматическом ретикулюме.
Запасы АТФ в мышце незначительны и, чтобы поддержать их деятельность, необходим непрерывный ресинтез АТФ. Он происходит за счет энергии, получаемой при распаде креатинфосфата (КрФ) на креатин (Кр) и фосфорную кислоту (анаэробная фаза). С помощью ферментов фосфатная группа от КрФ быстро переносится на АДФ (в течение тысячных долей секунды). При этом на каждый моль КрФ освобождается 46 кДж:
 АДФ + КрФ АТФ + Кр.
АДФ + КрФ АТФ + Кр.
Присоединение к креатину НзРО4 приводит к ресинтезу КрФ. При истощении запасов КрФ более медленный ресинтез АТФ осуществляется с помощью гликолетических и окислительных процессов. В анаэробных условиях необходимая энергия освобождается в процессе расщипления углеводов (гликогена и глюкозы). Под влиянием гликолитических ферментов они распадаются до молочной кислоты с выделением энергии. В результате гликолиза НзРО4 в начале присоединяется к углеводам, а затем, обогащенная энергией, отщепляется от них. В результате образуется богатая энергией фосфатная связь, которая и используется для ресинтеза АТФ.
Аэробный ресинтез АТФ (синонимы: окислительное фосфорилирование, тканевое дыхание) в 20 раз эффективнее анаэробного энергообразования. Накопленная во время анаэробной деятельности и в процессе длительной работы часть молочной кислоты окисляется до углекислоты и воды (1/4−1/6 ее часть), образующаяся энергия используется на восстановление оставшихся частей молочной кислоты в глюкозу и гликоген, при этом обеспечивается ресинтез АТФ и КрФ. Энергия окислительных процессов используется также и для ресинтеза углеводов, необходимых мышце для ее непосредственной деятельности.
В целом углеводы дают наибольшее количество энергии для мышечной работы. Например, при аэробном окислении глюкозы образуется 38 молекул АТФ (для сравнения: при анаэробном распаде углевода образуется лишь 2 молекулы АТФ).
Время развертывания аэробного пути образования АТФ составляет 3−4 мин (у тренированных до 1 мин), максимальная мощность при этом 350–450 кал/мин/кг, время поддержания максимальной мощности – десятки минут. Если в покое скорость аэробного ресинтеза АТФ невысокая, то при физических нагрузках его мощность становится максимальной и при этом аэробный путь может работать часами. Он отличается также высокой экономичностью: в ходе этого процесса идет глубокий распад исходных веществ до коноечных подуктов − СО2 и Н2О. Кроме того, аэробный путь ресинтеза АТФ отличается универсальностью в использовании субстратов: окисляются все органические вещества организма (аминокислоты, белки, углеводы, жирные кислоты, кетоновые тела и др.).
Однако аэробный способ ресинтеза АТФ имеет и недостатки:
1) он требует потребления кислорода, доставка которого в мышечную ткань обеспечивается дыхательной и сердечно-сосудистой системами, что, естественно, связано с их напряжением;
2) любые факторы, влияющие на состояние и свойство мембран митохондрий, нарушают образование АТФ;
3) развертывание аэробного образования АТФ продолжительно по времени и невелико по мощности.
Мышечная деятельность, осуществляемая в большинстве видов спорта, не может полностью быть обеспечена аэробным процессом ресинтеза АТФ, и организм вынужден дополнительно включать анаэробные способы образования АТФ, имеющие более короткое время развертывания и большую максимальную мощность процесса (то есть наибольшее количество АТФ, образуемое единицу времени), 1 моль АТФ соответствует 7,3 кал или 40 Дж (1 кал = 4,19 Дж).
Возвращаясь к анаэробным процессам энергообразования, следует уточнить, что они протекают по меньшей мере в виде двух реакций:
1. Креатинфосфокиназная, когда осуществляется расщепление КрФ, фосфорные группировки с которого переносятся на АДФ, ресинтезируя при этом АТФ. Но запасы креатинфосфата в мышцах невелики и это обусловливает быстрое (в течение 2−4 с) угасание этого типа реакции.
2. Гликолитическая (гликолиз) – развивается медленнее, в теченние 2−3 мин интенсивной работы. Гликолиз начинается с фосфолирирования запасов гликогена мышц и поступающей с кровью глюкозы. Энергии этого процесса хватает на несколько минут напряженной работы. На этом этапе завершается первая стадия фосфорилирования гликогена и происходит подготовка к окислительному процессу. Затем наступает вторая стадия гликолитической реакции – дегидрогенирование и третья – восстановление АДФ в АТФ. Гликолитическая реакция заканчивается образованием двух молекул молочной кислоты, после чего разворачиваются дыхательные процессы (к 3−5 минутам работы), когда начинает окисляться молочная кислота (лактат), образованная в процессе анаэробных реакций.
Биохимическими показателями оценки креатинфосфатного анаэробного пути ресинтеза АТФ являются креатининовый коэффицент и алактатный (без молочной кислоты) кислородный долг. Креатининовый коэффициент – это выделение креатинина с мочой за сутки в расчете на 1 кг массы тела. У мужчин выделение креатинина колеблется в пределах 18─32 мг/сут х кг, а у женщин – 10─25 мг/сут х кг. Между содержанием креатинфосфата и образованием у него креатинина существует прямолинейная зависимость. Следовательно, с помощью креатининового коэффициента можно оценить потенциальные возможности этого пути ресинтеза АТФ.
Биохимические сдвиги в организме, обусловленные накоплением молочной кислоты в результате гликолиза. Если в покое до начала мышечной деятельности концентрация лактата в крови составляет 1,2 ммоль/л, то после интенсивных непродолжительных нагрузок в течение 2─3 мин эта величина может достигать 18─20 ммоль/л. Другим признаком, отражающим накопление в крови молочной кислоты, служит показатель крови (рН): в покое ─ 7,36, после нагрузки снижение до 7,0 и более. Накопление лактата в крови определяет и ее щелочной резерв – щелочные компоненты всех буферных систем крови.
Окончание интенсивной мышечной деятельности сопровождается снижением потребления кислорода в начале резко, затем более плавно. В связи с этим выделяют два компонента кислородного долга: быстрый (алактатный) и медленный (лактатный). Лактатный – это то количество кислорода, которое используется после окончания работы для устранения молочной кислоты: меньшая часть окисляется до Н2О и СО2, большая часть превращается в гликоген. На это превращение тратится значительное количество АТФ, которая образуется аэробным путем за счет кислорода, составляющего лактатный долг. Метаболизм лактата осуществляется в клетках печени и миокарда.
Количество кислорода, необходимое для полного обеспечения выполняемой работы, называют кислородным запросом. Например, в беге на 400 м кислородный запрос равен приблизительно 27 л. Время пробегания дистанции на уровне мирового рекорда составляет около 40 с. Исследования показали, что за это время спортсмен поглощает 3─4 л О2. Следовательно, 24 л – это общий кислородный долг (около 90 % кислородного запроса), который ликвидируется после забега.
В беге на 100 м кислородный долг может доходить до 96 % запроса. В беге на 800 м доля анаэробных реакций несколько снижается ─ до 77 %, в беге на 10 000 м ─ до 10 %, то есть преобладающая часть энергии поставляется за счет дыхательных (аэробных) реакций.
Механизм мышечного расслабления.Как только в мышечное волокно перестают поступать нервные импульсы, ионы Са++ под действием так называемого кальциевого насоса за счет энергии АТФ уходят в цистерны саркоплазматического ретикулюма и их концентрация в саркоплазме понижается до исходного уровня. Это вызывает изменения конформации тропонина, который, фиксируя тропомиозин в определенном участке актиновых нитей, делает невозможным образование поперечных мостиков между толстыми и тонкими нитями. За счет упругих сил, возникающих при мышечном сокращении в коллагеновых нитях, окружающих мышечное волокно, оно при раслаблении возвращается в исходное состояние. Таким образом, процесс мышечного расслабления, или релаксации, так же как и процесс мышечного сокращения, осуществляется с использованием энергии гидролиза АТФ.
Входемышечнойдеятельностивмышцахпоочереднопроисходятпроцессысокращенияирасслабленияи, следовательно, скоростно-силовые качества мышц в равной мере зависят от скорости мышечного сокращения и способности мышц к релаксации.
Краткая характеристика гладких мышечных волокон.В гладких мышечных волокнах отсутствует миофибриллы. Тонкие нити (актиновые) соединены с сарколеммой, толстые (миозиновые) находятся внутри мышечных клеток. В гладких мышечных волокнах отсутствуют также цистерны с ионами Са++. Под действием нервного импульса ионы Са++ медленно поступают в саркоплазму из внеклеточной жидкости и также медленно уходят после того, как прекращают поступать нервные импульсы, поэтому гладкие мышечные волокна медленно сокращаются и медленно расслабляются.
Общий обзор скелетных мышц человека.Мышцытуловищавключаютмышцыгруднойклетки, спиныи живота. Мышцы грудной клетки участвуют в движениях верхних конечностей, а также обеспечивают произвольные и непроизвольные дыхательные движения. Дыхательные мышцы грудной клетки называются наружными и внутренними межреберными мышцами. К дыхательным мышцам относятся также и диафрагма. Мышцы спины состоят из поверхностных и глубоких мышц. Поверхностные обеспечивают глубокие, некоторые движения верхних конечностей головы и шеи («выпрямители туловища») прикрепляются к остистым отросткам позвонков и тянутся вдоль позвоночника. Мышцы спины участвуют в поддержании вертикального положения тела, при сильном напряжении (сокращении) вызывают прогибание туловища назад. Брюшные мышцы поддерживают давление внутри брюшной полости (брюшной пресс), участвуют в некоторых движениях тела (сгибание туловища вперед, наклоны и повороты в стороны), в процессе дыхания.
Мышцы головы и шеи – мимические, жевательные и приводящие в движение голову и шею. Мимические мышцы прикрепляются одним своим концом к кости, другим – к коже лица, некоторые могут начинаться и оканчиваться в коже. Мимические мышцы обеспечивают движения кожи лица, отражают различные психические состояния человека, сопутствуют речи и имеют значение в общении. Жевательные мышцы при сокращении вызывают движение нижней челюсти вперед и в стороны. Мышцы шеи участвуют в движениях головы. Задняя группа мышц, в том числе и мышцы затылка, при тоническом (от слова «тонус») сокращении удерживают голову в вертикальном положении.
Мышцы верхних конечностей обеспечивают движения плечевого пояса, предплечья и приводят в движение кисть и пальцы. Главными мышцами-антагонистами являются двуглавая (сгибатель) и трехглавая (разгибатель) мышцы плеча. Движения верхней конечности и, прежде всего, кисти чрезвычайно многообразны. Это связано с тем, что рука человека выполняет рабочую функцию.
Мышцы нижних конечностей обеспечивают движения бедра, голени и стопы. Мышцы бедра играют важную роль в поддержании вертикального положения тела, но у человека они развиты сильнее, чем у других позвоночных. Мышцы, осуществляющие движения голени, расположены на бедре (например, четырехглавая мышца, функцией которой является разгибание голени в коленном суставе; антагонист этой мышцы – двуглавая мышца бедра). Стопа и пальцы ног приводятся в движение мышцами, расположенными на голени и стопе. Сгибание пальцев стопы осуществляется при сокращении мышц, расположенных на подошве, а разгибание ─ мышцами передней поверхности голени и стопы. Многие мышцы бедра, голени и стопы принимают участие в поддержании тела человека в вертикальном положении.