Механизм и условия развития мышечного утомления Локализация утомления Концепции утомления
Вернемся к нашей задаче – рассмотрим механизмы развития физического утомления. Физическое утомление, которое является частным случаем утомления вообще, но основным видом утомления при занятиях физической культурой и спортом, связано, прежде всего, с развитием функциональных нарушений в ходе непосредственной мышечной деятельности. Внешние проявления, при развития такого утомления, будут связаны с функциональными нарушениями самого двигательного аппарата.
Вспомним, что функциональной единицей двигательного аппарата является нейромоторная (двигательная) единица – комплекс из мотонейрона (двигательного нейрона) и группы мышечных волокон, с которыми нейрон связан через синапс, и сокращение которых запускает. Для понимания механизма развития утомления следует выяснить меру участия каждой из этих структур, что можно сделать, поочерёдно исключая элементы двигательных единиц из сократительного процесса. Такими элементами будут: нервный центр, двигательный нейрон, нейромышечный (мионевральный) синапс и мышечные волокна.
При изучении сократительной деятельности мышцы не изолированной, а связанной с нервной системой, очевидным становится тот факт, что при работе двигательного аппарата утомление возникает раньше в нервных звеньях - структурах центральной нервной системы, затем в синаптических окончаниях нервных волокон двигательных нейронов - и в последнюю очередь в самой мышце. Причем, нервные волокна при этом характеризуются относительной неутомляемостью
Экспериментальным доказательством этого может служить следующий опыт.
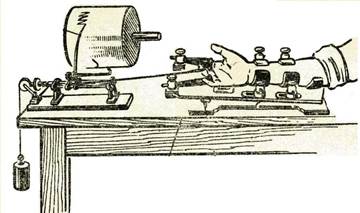 Если предложить человеку ритмически производить подъем груза мышцей (регистрируя результат например, на эргографе Моссо, рис.2), то через некоторое время возникает утомление – амплитуда мышечных сокращений начинает заметно снижаться. Однако, электрическое раздражение пороговой силы, приложенное к соответствующему двигательному нерву, на этом фоне, заставит мышцу сокращаться с прежней амплитудой. Это свидетельствует о том, что утомление возникает, прежде всего, в нервных центрах.
Если предложить человеку ритмически производить подъем груза мышцей (регистрируя результат например, на эргографе Моссо, рис.2), то через некоторое время возникает утомление – амплитуда мышечных сокращений начинает заметно снижаться. Однако, электрическое раздражение пороговой силы, приложенное к соответствующему двигательному нерву, на этом фоне, заставит мышцу сокращаться с прежней амплитудой. Это свидетельствует о том, что утомление возникает, прежде всего, в нервных центрах.
Через некоторое время мышца перестает отвечать и на раздражение, приложенное к двигательному нерву. Вместе с тем в ответ на электрическое раздражение, приложенное к самой мышце, она способна некоторое время сокращаться с прежней амплитудой. Следовательно, вторым звеном, в котором возникает процесс утомления, являются окончания двигательного нерва - мионевральная (синаптическая) передача.
Рис 2. Регистрация сокращений на эргографе Моссо.
Следовательно, мышца утомляется в последнюю очередь. При этом, имеет место очень сложный комплекс изменений как в возбудимой системе мышцы, (что находит свое выражение в уменьшении интенсивности токов действия и увеличении их продолжительности, заметном уменьшении скорости распространения возбуждения), так и, в сократительной способности ее двигательных единиц, в миофибриллах и других белковых системах обеспечивающих сокращение, а также в ходе энергетических процессов, обусловливающих активность миофибрилл.
В более простом варианте пронаблюдать роль каждого из этих элементов в развитии утомления можно и на нервно-мышечном препарате лягушки, но в этом случае невозможно оценить роль центрального звена. Зато на нервно-мышечном препарате можно изучить развитие утомления в зависимости от характера сократительой деятельности. Так, если утомлять мышцу, подвергая ее длительному ряду одиночных раздражений, то с течением времени амплитуда одиночных сокращений падает, длительность каждого сокращения нарастает, степень расслабления уменьшается, накапливается остаточное сокращение - контрактура. Тетаническое сокращение мышцы при продолжительной деятельности изменяется следующим образом: амплитуда сокращения постепенно уменьшается до полного прекращения; в момент утомления сплошной тетанус, особенно при условии слабых тетанизирующих раздражителей, становится зубчатым. При утомлении после прекращения тетанизирующего раздражения всегда следует контрактурное последствие.
В целостном организме, при нормальном кровоснабжении, утомление в мышце возникает позже, чем в условиях сосудистой изоляции мышц. При нормальном функционировании центральных и периферических аппаратов мышца работает более или менее длительное время без утомления. Субмаксимальные сокращения и сравнительно небольшие (оптимальные) нагрузки, являются теми наиболее благоприятными условиями, при которых мышца может работать длительно, и утомление не наступает в течение многих часов.
В условиях динамической нагрузки, при работе, лежащей ниже предела утомления, во время рабочих движений за счет достаточного времени расслабления мышц макроэргические фосфаты, используемые при сокращении, могут регенерировать, а конечные продукты обмена – удаляться. Время расслабления соответствует необходимому времени восстановления. Так как в этом случае остаточные явления утомления отсутствуют, такая работа называется не утомительной. При динамической работе, лежащей выше предела утомления, возможность непрерывного восстановления отсутствует, так как длительность периодов расслабления меньше, чем время, необходимое для восстановления. Восстановление запасов энергии и удаление молочной кислоты происходят не полностью и возникает накопление остаточного утомления. Мышца использует свои макроэргические субстраты и накапливает конечные продукты метаболизма, причем утомление нарастает. Выраженность мышечного утомления при динамической работе, лежащей выше пределов утомления, может быть определена на основании физиологических показателей (например, времени восстановления, пульсовой суммы восстановления; см. выше).
Утомление при статической работе.В этом случае мышечное утомление вызвано в основном неадекватным кровотоком. Если сила мышечного сокращения превышает 15%изометрического максимума, приток кислорода перестает соответствовать потребности в нем и мышечное утомление прогрессивно нарастает.
Современные концепции утомления складываются из представлений о многоструктурности и неоднозначности функциональных изменений в отдельных системах во время работы. В зависимости от вида работы, ее напряженности, продолжительности ведущая роль в развитии утомления может принадлежать различным физиологическим системам.
Изменения в гуморальной системе регуляции могут стать ведущими факторами утомления при напряженной мышечной работе, связанной с эмоциональным стрессом.
При длительной истощающей работе наряду с предельными затратами энергии продолжение работы может лимитировать и утомление системы гипоталамус – гипофиз – надпочечники.
Нарушения в центральном звене регуляции физиологических функций могут играть существенную роль в развитии утомления при кратковременной мышечной работе скоростного характера. В результате мощного потока проприоцептивных и хеморецептивных импульсов в ЦНС развивается запредельное торможение (первичное утомление). Чрезмерная частота нервных импульсов к исполнительным приборам истощает и генерирующие их нервные клетки. Уже через несколько секунд работы падает лабильность нервных центров, в результате чего снижается и скорость выполнения упражнений.
Снижение скорости ресинтеза АТФ вследствие накопления продуктов промежуточного обмена может рассматриваться как главный фактор, ограничивающий продолжительность интенсивной работы. В скелетных мышцах поддерживается относительно постоянная концентрация АТФ, расходование ее инициирует компенсаторные процессы: повышается активность окислительных ферментов. Углеводы, свободные жирные кислоты и аминокислоты окисляются в митохондриях. При этом освобождается энергия, которая идет на ресинтез АТФ или запасается в макроэргических связях креатинфосфата (КрФ). При работе в анаэробных условиях ресинтез АТФ идет с накоплением молочной кислоты.
Переключение на анаэробные источники энергии при работе определяется не только ее интенсивностью, но и уровнем тренированности спортсмена. Чем ниже этот уровень, тем быстрее совершается переход на менее экономичный способ получения энергии, тем быстрее развивается некомпенсируемое утомление. Избыток молочной кислоты в мышцах может приводить к разобщению процессов образования энергии в окислительном цикле и накоплению ее в фосфагенах – АТФ и КрФ. Поэтому спортсмен с невысоким уровнем тренированности отказывается от работы значительно раньше, чем истощаются энергетические ресурсы. Молочная кислота служит источником водородных ионов. Их избыток в сократительном аппарате мышц препятствует образованию актомиозиновых мостиков, само сокращение мышцы затрудняется.
Подключение гликолиза к энергообеспечению происходит при высокой мощности работы через 20-30с после ее начала. Накапливающиеся при этом продукты обмена угнетают лилолитические процессы, тормозят окислительное фосфорилирование. Накопление лактата вмышцах является, по-видимому, главным фактором развития утомления при работе субмаксимальной мощности,
При работе большой мощности главной причиной развития утомления является относительная гипоксия ткани, а также постепенное накопление продуктов гликолиза и их угнетающее действие на аэробный обмен в мышцах, на процессы нервной регуляции двигательной функции. Парадоксальное на первый взгляд развитие гипоксии в условиях предельного потребления кислорода и переход на использование энергии гликолиза объясняется тем, что потребность в кислороде при работе большой мощности намного выше максимально возможного его потребления. Часть энергии организм вынужден черпать из анаэробного расщепления глюкозы – гликолиза. Отсюда избыток молочной кислоты.
При работе умеренной мощности на первое место в развитии утомления выходит истощение энергетических ресурсов – главным образом гликогена в печени и в работающих мышцах. Нарушения в регуляторных влияниях ЦНС являются, вероятно, вторичными.
Сдвиги в химизме внутренней среды в результате накопления продуктов промежуточного обмена отражаются в первую очередь на состоянии функций высших корковых и подкорковых регуляторов физиологических функций. Образуется порочный круг нарушения регуляторных механизмов. Первичные метаболические расстройства усугубляются нарушением регуляторных влияний со стороны ЦНС.
Строгая количественная оценка значимости отдельных факторов в развитии утомления при конкретных видах мышечной работы является одним из важных элементов управления тренировочным процессом. Выделение ведущего фактора возможно при правильном подборе тестирующих проб и методик исследования.
Утомление у детей школьного возраста развивается быстрее, чем у взрослых, вследствие ряда особенностей деятельности ЦНС. У детей быстрее нарушаются процессы внутреннего торможения, в особенности дифференцировочного и запаздывающего. При этом ухудшается внимание, появляется двигательное беспокойство, сменяющееся резким спадом активности в результате развития охранительного торможения и понижения возбудимости корковых клеток, Дети отказываются от работы задолго до развития критического состояния, связанного с накоплением продуктов промежуточного распада и тем более истощением энергетических источников.
В занятия с детьми не следует включать однообразную, монотонную работу. Необходимо чередовать различные виды работы, облегчающие восстановление по механизму активного отдыха. Учитывая, что работоспособность у детей падает от первого урока к последнему, от понедельника ксубботе, в режиме дня следует предусмотреть соответствующее содержание физических и умственных нагрузок, а также средства и методы их активизации.
Позно-тонические реакции
Двигательная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы.
В основе двигательной деятельности человека лежат 2 формы механической реакции мышечных волокон: 1) длительное тоническое напряжение и 2). фазное (тетаническое) сокращение
Фазная деятельность обеспечивает быстрые и сильные сокращения мышц при выполнении движений, а также коррекции позы. Эти сокращения осуществляются быстрыми двигательными единицами.
Тоническое напряжение отличается от фазного медленным развитием, большей экономичностью, преобладанием изометрического режима и преимущественным участием медленных двигательных единиц. Оно лежит в основе поддержания определенной позы тела. Если при поддержании позы требуется значительное усилие мышц (например для выполнении угла в упоре), то включаются фазные сокращения мышц.
Рефлекторная природа мышечного тонуса. В поддержании тонического напряжения мышц участвует сложная многоуровневая система нервных центров, находящаяся под контролем коры. больших полушарий.
По своей природе мышечный тонус является рефлекторным актом. Для его возникновения в мышцах достаточна уже рефлекторная деятельность спинного мозга. Однако тонкое перераспределение тонуса между различными мышцами, необходимое для протекания реакций целостного организма, осуществляется более высокими этажами центральной нервной системы, а его произвольная регуляция — корой больших полушарий.
В наиболее простом случае появление тонуса обусловлено сокращением мышцы в ответ на раздражение проприорецепторов при ее растяжении. Такой рефлекс, осуществляемый спинным мозгом, называется миотатическим рефлексом на растяжение. Если это растяжение длительно, то рефлекторное сокращение носит тоже длительный тонический характер.
В естественных условиях основным раздражителем рецепторов мышечных и сухожильных веретен является сила тяжести, растягивающая скелетные мышцы, особенно мышцы-разгибатели. Поражение спинного мозга приводит к исчезновению тонуса мышц.
 Гамма-регуляция мышечного тонуса. Степень тонического напряжения мышцы зависит так же от частоты импульсов, посылаемых к ней тоническими альфа-мотонейронами. Частота разрядов альфа-мотонейронов, в свою очередь, регулируется импульсами от проприорецепторов той же самой мышцы. Получается замкнутое кольцо между мышцей и иннервирующими ее мотонейронами. Однако потоки импульсов в этом кольце могут регулироваться вышележащими этажами нервной системы с помощью гамма-мотонейронов спинного мозга (рис.1). Разряд гамма-мотонейронов повышает чувствительность мышечных веретен. В результате увеличивается поток импульсов от рецепторов к альфа-мотонейронам и от альфа-мотонейронов к мышце. Тем самым повышается мышечный тонус.
Гамма-регуляция мышечного тонуса. Степень тонического напряжения мышцы зависит так же от частоты импульсов, посылаемых к ней тоническими альфа-мотонейронами. Частота разрядов альфа-мотонейронов, в свою очередь, регулируется импульсами от проприорецепторов той же самой мышцы. Получается замкнутое кольцо между мышцей и иннервирующими ее мотонейронами. Однако потоки импульсов в этом кольце могут регулироваться вышележащими этажами нервной системы с помощью гамма-мотонейронов спинного мозга (рис.1). Разряд гамма-мотонейронов повышает чувствительность мышечных веретен. В результате увеличивается поток импульсов от рецепторов к альфа-мотонейронам и от альфа-мотонейронов к мышце. Тем самым повышается мышечный тонус.
Изменение напряжения мышц под влиянием деятельности гамма-мотонейронов называется гамма-регуляцией. Активность гамма-мотонейронов находится под контролем ретикулярной формации ствола мозга. В регуляции их деятельности, а следовательно, и в регуляции тонуса скелетных мышц участвуют мозжечок, подкорковые и другие структуры экстрапирамидной системы. Высший контроль осуществляется корой больших полушарий. Рис 1. Гамма-регуляция деятельности мышц
Значение различных отделов головного мозга в регуляции тонуса скелетных мышц. Различные отделы головного мозга, посылая импульсы к вставочным нейронам и мотонейронам спинного мозга, могут регулировать тонус скелетных мышц, изменяя тем самым позные и двигательные реакции организма. Влияния на мышечный тонус оказывают обе системы головного мозга — как специфическая, так и неспецифическая.
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел среднего мозга, а угнетение – тормозящий отдел продолговатого мозга.
Специфические системы действуют избирательно – на отдельные группы мышц. Усиление тонуса мышц-сгибателей вызывают кортико-спинальная, кортико-рубро-спинальная и частично кортико-ретикуло-спинальная системы (последняя оказывает также неспецифическое диффузное влияние). Одновременно эти системы снижают тонус мышц-разгибателей. В противоположность им вестибуло-спинальная система повышает тонус мышц-разгибателей и тормозит тоническое напряжение мышц-сгибателей. Изменения тонуса могут происходить очень быстро, нередко опережая двигательные акты. Это позволяет организму подготовиться к необходимому движению, заранее изменив позу тела.
Позы. Позой называется закрепление частей скелета в определенном положении.
Организация позы необходима для преодоления силы земного притяжения. Поза человека служит для сохранения равновесия тела в состоянии двигательного покоя и при выполнении статической и динамической работы. Движения человека всегда происходят на фоне определенной позы тела.
Поддержание позы осуществляется как длительными и экономичными тоническими напряжениями мышц, так и быстрыми фазными сокращениями при больших нагрузках и в моменты коррекции позы. Часто позно-тонические реакции мышц бывают кратковременными, так как возникают только в определенные фазы движений (например, фиксация суставов при отталкивании от опоры).
Поза, принимаемая при работе, называется рабочей. Рациональная ее организация снижает утомление мышц и повышает работоспособность человека. Большое значение имеют предрабочие изменения позы, которые заранее компенсируют возможные изменения центра тяжести тела и предотвращают его падение, а также препятствуют действию реактивных сил. Для отдыха и во время выполнения работы человек выбирает наиболее удобную позу, при которой возможно минимальное напряжение скелетных мышц.
Частным случаем позных реакций является поддержание заданного усилия мышцы и заданного суставного угла.
Основной позой человека является вертикальное положение тела теменем кверху – поза стояния. Поддержание этой позы в основном обеспечивается тоническим напряжением мышц-разгибателей задней поверхности тела, составляющих так называемый антигравитационный аппарат, Положение равновесия в этой позе неустойчиво из-за небольшой площади опоры и высокого положения центра тяжести. Тело человека все время совершает небольшие колебания, компенсируемые рефлекторными сокращениями антигравитационных мышц,
Поза сиденияявляется более удобной, так как характеризуется дополнительной опорой на седалищные бугры (около 50% веса тела). В течение нескольких минут естественная поза сидения поддерживается без заметного напряжения спинных мышц, активность которых нарастает при утомлении.
Поза лежания характеризуется наименьшей активностью мышц, которые в наибольшей степени освобождаются от активного поддержания или фиксации частей тела.
В спортивной деятельности встречаются позы с необычным положением тела – висы, упоры, стойки. Они требуют длительной тренировки дляих освоения, формирования специализированных навыков.
Сохранению позы способствуют установочные рефлексы. К ним относятся статические и статокинетические рефлексы, в осуществлении которых большое значение имеет продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве:
1) при изменении положения головы в пространстве – это так называемые лабиринтные рефлексы. возникающие в результате раздражения рецепторов вестибулярного аппарата;
2) при изменении положения головы по отношению к туловищу – шейные рефлексы, с проприорецепторов мышц шеи
3) при нарушении нормальной позы тела – выпрямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед – тонус сгибателей (лабиринтный рефлекс). Подобные реакции имеют место при выполнении многих физических упражнений. Так, выполнение гимнастом стойки на кистях облегчается при отклонении головы назад выполнение группировки во время прыжков в воду или акробатических прыжков – при наклоне головы вперед. Выпрямительные рефлексы – это последовательные сокращения мышц шеи и туловища которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У человека они проявляются, например, во время ныряния.
Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется – это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение – нистагм глаз – обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
В двигательной деятельности человека часто возникают ситуации, когда требуется подавить эти установочные рефлексы. Произвольное подавление врожденных установочных рефлексов продолговатого и среднего мозга обеспечивается тормозящими влияниями из коры полушарий. Например, для спринтера невыгодно раннее выпрямление туловища при стартовом разбеге, поэтому корой больших полушарий тормозится выпрямительный рефлекс
Двигательная деятельность
Движения человека являются комплексом различных двигательных актов, которые обеспечиваются как сравнительно простыми двигательными рефлексами, так и сложнейшими их комбинациями.
Двигательные акты различной сложности. Рассматривая различные двигательные акты человека, можно выделить
- элементарные двигательные рефлексы,
- более сложные – ритмические рефлексы и, наконец,
- особенно сложные формы двигательной деятельности, обеспечивающие поведение человека.
Элементарные двигательные рефлексы. При сложных движениях человека используются элементарные двигательные рефлексы, осуществляемые спинным мозгом. К простым безусловным двигательным рефлексам спинного мозга относятся:
- рефлексы, на растяжение (миотатические и сухожильные рефлексы),
- сгибательные рефлексы на раздражение кожных рецепторов (рефлексы удаления от раздражителя)
- рефлексы отталкивания (рефлексы сближения с опорой).
Миотатические и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий. Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению. Рефлекторная дуга рефлекса на растяжение – моносинаптическая. Простота рефлекторной дуги обеспечивает быструю и точную реакцию мышцы, меньшую утомляемость рефлекторных ответов.
Рефлексы на растяжение имеют большое значение при статической деятельности мышц для поддержания определенной позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В произвольной двигательной деятельности человека иногда необходимо подавлять эти рефлексы (например, выполнение гимнастического шпагата требует подавления рефлекса на растяжение).
При раздражении рецепторов кожи (болевых, температурных и др.) возникает сокращение мышц-сгибателей, которое позволяет отдернуть конечность от повреждающего раздражителя. Этот сгибательный кожный рефлекс носит защитный характер. В его основе лежит полисинаптическая рефлекторная дуга. В отличие от моносинаптического рефлекса на растяжение при сгибательном рефлексе быстрее наступает утомление; время этого рефлекса в связи с наличием вставочных нейронов значительно больше.
Одна из разновидностей элементарных рефлексов спинного мозга – рефлекс отталкивания, описанный Ч. Шеррингтоном. Этот рефлекс возникает при раздражении кожи стопы опорой. Однако, в Отличие от сгибательного рефлекса, он приводит не к отдергиванию конечности от раздражителя, а к сближению с раздражителем. В первом случае мы имеем защитный рефлекс, предохраняющий организм от разрушения. Во втором случае – биологически целесообразный акт, обеспечивающий контакт с опорой при стоянии и отталкивание от нее при передвижениях. Этот рефлекс лежит в основе сложных локомоций: ходьбы, бега, прыжков в длину ил' высоту и др.
Ритмические рефлексы. Составной частью различных сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Они особенно выражены при выполнении циклических движений. Их возникновение и протекание связаны с проявлением механизмов взаимосочетанной (реципрокной) иннервации мышц-антагонистов, при которой возбуждение центров-сгибателей сопровождается одновременным торможением центров мышц-разгибателей. При этом проявляется и, так называемый, рефлекс отдачи: после акта сокращения наступает расслабление, а затем снова акт сокращения. Рефлекс отдачи – это рефлекс на растяжение с проприорецепторов мышц и сухожилий, в результате которого возникает сокращение растянутого разгибателя. Такая простая форма ритмического рефлекса часто встречается в двигательной деятельности человека (например, при забивании молотком гвоздей).
Эта форма рефлексов – одна из древних и относительно простых форм – имеет большое значение в организации многих сложных движений. Например, включение ритмических циклоидных движений в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи; то же самое происходит при освоении акта ходьбы – переход от отдельных шагов к ритмической походке (Н. А. Бернштейн).
Наиболее простая форма ритмического рефлекса – чесательный рефлекс животных, который обеспечивается попеременным сокращением и расслаблением одних и тех же мышц одной конечности. Более сложная форма – шагательный рефлекс, лежащий в основе ходьбы, бега и других локомоций. Подключение второй конечности к координированному двигательному акту происходит в результате осуществления перекрестного рефлекса. Этот рефлекс осуществляется с участием вставочных тормозных нейронов (клеток Рэншоу), вызывающих торможение мотонейронов на противоположной стороне спинного мозга. В целостном организме человека сгибание одной ноги вызывает перекрестный разгибательный рефлекс другой ноги, принимающей на себя тяжесть тела при стоянии, ходьбе и пр.
Включение этого спинального механизма производится с более высоких этажей нервной системы. В так называемой «локомоторной области» среднего мозга имеются центры, электрическое раздражение которых запускает шагательные движения конечностей.
В осуществлении шагательного рефлекса принимает участие и мозжечок. Удаление одного его полушария у животных приводит к искажению движений (особенно передней конечности). Движения сильно варьируют по амплитуде, заметно искажаются при малейших посторонних воздействиях, происходит чрезмерное сгибание локтевого сустава в фазе переноса конечности.
Протекание ритмических рефлексов, в том числе шагательного, связано с деятельностью различных отделов неспецифической системы. При активирующих влияниях происходит нарастание амплитуды движений, а при угнетающих влияниях – ее снижение.
Важное значение, в регуляции ритмических рефлексов, имеют подкорковые ядра – бледное ядро и полосатое тело, обеспечивающие их автоматическое протекание, содружественные движения конечностей, вспомогательные реакции (фиксацию суставов и др.).
Высшим регулятором рефлексов является кора больших полушарий, особенно ее премоторная область. Благодаря коре ритмические движения (например, простой акт ходьбы) приобретают определенное смысловое значение, включаются как составной элемент сложные акты поведения.
Сложные формы двигательной деятельности. В целостном поведении простые рефлексы, сочетаясь, обусловливают сложные двигательные действия. Социальные условия жизни человека намного усложняют его двигательную деятельность, приводя к появлению специально человеческих форм движений: бытовых, производственных, спортивных. Простые и сложные ритмические рефлексы лежат в основе циклической двигательной деятельности человека: ходьбы, бега, плавания, гребли, ходьбы на лыжах, езды на велосипеде и пр. Большое значение в их осуществлении имеют механизмы реципрокной иннервации мышц-антагонистов.
Однако в целом ряде движений механизмов реципрокной иннервации мышц-антагонистов становится недостаточно, требуется одноименная работа мышц-антагонистов – в одной конечности или даже в обеих. В этих случаях реципрокные отношения, характерные для спинного мозга, подавляются центрами головного мозга. Особенно часто встречаются симметричные (а не реципрокно-перекрестные) движения в деятельности верхних конечностей человека, т. е. происходит одновременное сокращение одноименных мышц левой и правой рук.
Наиболее сложные формы движений представляют собой многофазную цепь рефлексов, которая основана уже не на элементарных реципрокных отношениях, а является целостным двигательным навыком, образующимся в процессе обучения по механизму условных рефлексов.