КАК И ПОЧЕМУ ВОЗНИКЛО СОЗНАНИЕ
Oacute; 2008 г. В.М.Верхлютов
Институт высшей нервной деятельности и нейрофизиологии РАН, Москва
e-mail: verkhliutov@mail.ru
Активирующая система является глобальным выключателем сознания. Основные части системы активации - ретикулярная формация, таламус, кора и белое вещество. Активация необходима для поддержания нормальной работы пирамидных нейронов коры испытывающих недостаток афферентных воздействий. Возникновение активирующей системы обусловлено случайной мутацией приведшей к избытку вставочных нейронов и дефициту афферентации. Эволюция активирующей системы сопровождалось возникновением феномена сознания, который основан на принципах самоактивации и внутреннего отражения. Сознание и память обеспечиваются единым нейрофизиологическим механизмом.
Ключевые слова: сознание, память, искусственный интеллект.
Феномен сознания - самый важный и самый загадочный из всех свойств психики, которая в свою очередь является обратной стороной функционирования мозга. Автор этих строк еще совсем недавно не подозревал, что приблизится к пониманию этого парадоксального явления. Но однажды его спросили: «Почему человек теряет сознание при ударе по голове, во время сна или находясь в коме» ?
При общении с любым сложным устройством первое, что необходимо понять, как его включать и выключать. Так и в нашем случае, надо найти выключатель сознания. Что поддерживает наше сознание в состоянии бодрствования? За ответом не пришлось далеко ходить. В 1949 году Джузеппе Моруцци и Хоакин Мегун опубликовали знаменитую работу [14], где было определено понятие активирующей системы и описана ретикулярная формация (РФ) ствола головного мозга.
Данной анатомической структуре приписывались самые невероятные качества, вплоть до морфологического субстрата человеческой души. В действительности она выполняет очень важную, но более скромную роль глобального выключателя для нейронов коры головного мозга. Самыми крупными и наиболее интересными из этих клеток мозга являются большие пирамиды, которые имеют большое количество контактов (синапсов) с одной стороны и недостаточно обеспечены афферентными входами с другой. Для их нормального функционирования необходимы обязательные и достаточные импульсные посылки от активирующих структур. Без активирующих воздействий обработка информации этими нейронами невозможна (рис.1).
Таким образом, необходимо разделить собственно информационные процессы в мозге и активирующие воздействия, выполняющие функции управления большим числом нейронов (рис 1). При этом нельзя исключить вариант, когда информационные каналы используются как активирующие и наоборот. Это позволяет пирамидным нейронам быть вычислительным элементом с большим динамическим диапазоном.
Если для обеспечения информационной операции данному нейрону необходимо небольшое количество контактов, то активирующие возбуждающие воздействия должны быть максимальны. При востребовании всех ресурсов данного нейрона информационным процессом активация минимальна.
Активирующие воздействия могут быть как возбуждающими, так и тормозными (рис.1). Возбуждающие и тормозные активирующие воздействия находятся в динамическом равновесии, хотя суммарно в нормально функционирующем мозге деполяризация (возбуждение) более выражена, чем гиперполяризация (торможение) в состоянии бодрствования, по сравнению с бессознательным состоянием или состоянием сна.
Во время сна фоновая деполяризация большинства пирамидных нейронов снижается на 10-15 мкв [5], что обусловлено снижением возбуждающих влияний активирующей системы. Исключением является фаза быстрого сна, где отмечается относительный подъем процессов деполяризации в сравнении с медленным сном. От бодрствования быстрый сон отличается тем, что сознание в этом случае оказывается изолированным от большинства внешних воздействий.
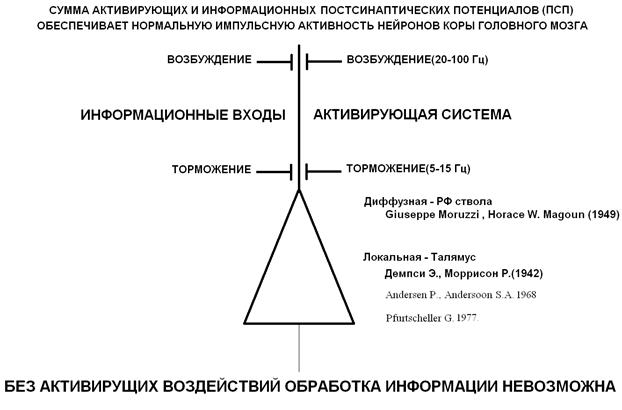
Рис 1. Активирующая и информационная системы мозга. Во время сна деполяризация пирамидных нейронов на 10-15 мкв меньше, чем во время бодрствования, что связано с уменьшением активности РФ, описанной Моруцци и Мегуном. Пробуждение сопровождается появлением альфа - ритма, механизм которого обусловлен талямо - кортикальными связями и описан Андерсенами. В состоянии бодрствования активация связана со снижением амплитуды альфа - ритма и появлений более быстрых бета и гамма – ритмов. Локальная десинхронизация подробно исследована Пфуртшилером.
Функция быстрого сна до конца еще не ясна, и возможно его значение чисто утилитарно. Например, запуск сознания во время сна необходим для равномерного распределения медиаторов в коре, которые поступают в мозг во время фазы медленного сна из депо в продолговатом мозге.
Ретикулярная формация не является единственной структурой, обеспечивающей работу активирующей системы. Один из признаков включения сознания - это появление на электроэнцефалограмме (ЭЭГ) альфа - ритма. Но, как ни странно, альфа – ритм отражает процессы торможения в коре. Механизм возникновения альфа – ритма обеспечивается прямыми и обратными таламо - кортикальными связями и был описан Андерсенами в 1968 г. (рис.1) [9].
В 1942 г. Демпси и Морисон показали обширные связи таламуса со всеми отделами коры [10].
Большинство ретикулярных путей продолговатого мозга оканчиваются в неспецифических таламическихя ядрах за исключением некоторой порции, которая непосредственно связывается с нейронами лобной коры и обеспечивает реакции пробуждения и внимания.
В состоянии спокойного бодрствования с закрытыми глазами альфа-ритм имеет максимальную амплитуду. В экспериментах с использованием функциональной магнитно-резонансной томографии (фМРТ) показано, что максимальное усиление альфа - ритма совпадает с повышением метаболизма в таламусе и его снижением в зрительной коре [11]. Наиболее мощный источник данного ритма локализован с помощью ЭЭГ и магнитоэнцефалограммы(МЭГ) [1,2] в зрительной коре. Альфа-ритм является оперативным тормозом обширных областей коры, и его усиление, при некоторых видах деятельности мозга, есть механизм вытормаживания функционально избыточных областей коры. (Другая гипотеза рассматривает альфа-ритм, как ритм замещения, выполняющий функцию замещения в при отсутствии сенсорного притока, что позволяет поддерживать уровень активации в нейросети на определенном уровне).
Эффект локальной десинхронизации альфа – ритма, привязанный к сигналам разной модальности, был открыт 1977 году Пфуртшилером [15] и в дальнейшем им подробно изучен [16]. Этим исследователем доказано, что локальная депрессия альфа - ритма сопровождается локальным возбуждением коры.
Физиологический источник альфа – ритма в отличие от физического дипольного источника находится в таламусе, который является вторым элементом активирующей системы, обеспечивающим управление мозга в состоянии сна и бодрствования.
Однако активирующие воздействия ствола и таламуса абсолютно недостаточны для обеспечения нормальной работы коры. Третьим элементом этой системы являются сами корковые нейроны (рис 2.). В коре даже простых животных были найдены нейроны – пейсмекеры для работы которых отсутствует необходимость сенсорного притока. В данном случае пейсмекер является аналогом сенсорного раздражителя с одной стороны, а с другой – выступает, как элемент системы активации.
Если ретикулярная формация аккумулирует возбуждение от сенсорных входов, таламус и кора способны самоактивироваться без каких либо внешних воздействий.
Функционирование коры и поддержание сознания обеспечивается тормозными и возбуждающими активирующими воздействиями ретикулярной формации, таламуса, коры и информационно оформленным потоком со стороны тех же образований (рис.2).
Для работы такого механизма необходима высокая степень синергии структур, которая реализуется посредством связей через белое вещество (аксоны). Повреждение белого вещества приводит к разной степени нарушения сознания, вплоть до полной его утраты в виде вегетативного состояния. Вегетативное состояние необходимо отличать от комы, как острого заболевания, сопровождающегося гипоксией и отеком мозга, и от состояния минимального сознания [17], при котором в основном нарушены внешние проявления сознания (отсутствуют или минимально выражены движения и речь) за счет дефицита влияний активирующей системы.
Можно предполагать, что различные виды нарушений сознания, включая изменения при психических расстройствах, связаны с дисфункцией активирующей системы. В недавних работах показан количественный дефицит и изменения миелинизации белого вещества при шизофрении, аутизме и т.д. [8].

Рис 2. Работа коры головного мозга с сенсорным потоком возможна только при включении активирующей системы, в состав которой входит ретикулярная формация, таламус и различные в эволюционном плане отделы коры. В информационном процессе, высшим проявлением которого является сознание, участвуют другие (специфические) отделы тех же структур. Белое вещество – аксоны ( обозначены на рисунке стрелками, где + и – есть возбуждающие и тормозные воздействия) являются важнейшим элементом этого механизма, обеспечивающего реализацию алгоритмов сознания.
Сознание - эволюционно целесообразное явление, позволяющее максимально приспособиться к условиям среды. Человеческое сознание, как наиболее развитое, позволило homo sapiens стать доминирующим видом на всей планете и выйти за ее пределы. Несмотря на свою исключительность, феномен сознания возник естественным путем, являясь логичным завершением эволюционного процесса усложнения и совершенствования нервной системы.
Строительство нервной системы началось с возникновения специализированных клеток – нейронов. Главным свойством нейрона является его возбудимость. По свойству возбудимости классифицируем нейроны на три группы: 1) нейроны-рецепторы, которые точно воспроизводят поступивший на вход сигнал, 2) нейроны-фильтры, преобразующие и видоизменяющие сигнал, 3) нейроны - пейсмекеры, обладающие нулевым или очень низким порогом возбудимости, и составляющие основу активирующей системы (рис.3). Нетрудно догадаться, что пейсмекеры появились в результате повышения чувствительности нейронов к афферентным сигналам в условиях необходимости реагировать на очень слабые внешние воздействия.
Возникновение активирующей системы в свою очередь было следствием дефицита афферентных воздействий, возникшего при случайной мутации, которая привела к избытку вставочных нейронов, необеспеченных достаточным информационным потоком.
Ретикулярная формация является самой древней структурой активирующей системы, и поэтому она сильно зависит от воздействий внешней среды. Таламус - эволюционно промежуточная структура, которая может работать автономно, независимо от внешних сигналов. Такими же свойствами обладает филогенетическисамая молодая структура мозга – кора, имеющая большое число нейронов - пейсмекеров и обладающая широкими возможностями самоактивации, т.к. восходящих активирующих воздействий абсолютно недостаточно для нормальной работы больших пирамидных нейронов (рис 3).
Именно с внутрикорковой активации началось формирование нейронных сетей, которые породили феномен сознания. Алгоритм сознания основан на взаимном анализе информации пирамидных нейронов друг от друга или двух нейронных сетей, которые отражают друг-друга.

Рис 3. Эволюция мозга шла по пути увеличения вставочных нейронов, что привело к дефициту рецепторов, появлению нейронов – пейсмекеров и формированию активирующей системы. Дальнейший прогресс был связан с дефицитом активирующих воздействий, что обусловило возникновение коллатеральной и дистантной активации внутри коры. Совершенствование этого вида активации завершилось появлением нейронов-фильтров и возникновением феномена сознания.
Нами получены экспериментальные данные, иллюстрирующие попеременную активацию полушарий в интервале от 350 до 1000 мсек после стимула со средней частотой 47 Гц [3]. Данный интервал, как было показано ранее другими исследователями, связан с загрузкой рабочей памяти [19], а частота соответствует бета- и гамма-ритмам. Попеременная активация полушарий происходила со средней частотой в диапазоне бета – ритма. В свете этих фактов, становится понятным наличие мощных связей между полушариями, интенсивно обменивающимися информацией. Одна из причин такого обмена - удержание недавно полученной информации для её анализа. Многократное повторение, предъявление одинаковых информационных паттернов на одни и те же входы при определенных условиях могут способствовать фиксации данной информации, что может быть основным механизмом долговременной памяти. Таким образом, процесс взаимного отражения информации пирамидными нейронами вероятно объединяет сознание и память.
Однако известны случаи однополушарных пациентов с приемлемым когнитивным дефицитом без критических нарушений сознаний. Это значит, что межполушарное отражение представляет собой только частный случай, и связи, обеспечивающие взаимное отражение, повсеместно распространены внутри полушария (рис.4). Экспериментальные данные подтверждают это предположение.
Показано, что в экстрастриарных областях имеются компактные участки с высокой концентрацией нейронов, реагирующих на определенную категорию стимулов [11]. При этом, в каждом полушарии имеется не менее двух подобных областей, расположенных симметрично относительно центральной линии ретинотопической проекции. Симметричные участки, которые можно обозначить как «категориальные модули», позволяют организовать взаимное отражение обрабатываемой и сохраняемой информации. Полученные нами экспериментальные данные позволяют предположить, что существуют взаимно отражающие связи не только между симметричными участками модуля, но и ближние связи внутри части категориального модуля [3].
Другие данные говорят о том, что обработка входного сигнала в мозге только отчасти последовательна. Это находит свое отражение в постепенном развертывании вызванного потенциала (ВП). Локализация компонентов ВП показала для каждого компонента наличие нескольких дипольных источников, которые остаются активными в той или иной степени в течении всего периода восприятия и обработки стимула [13]. Т.е., можно предположить наличие взаимного отражения сигналов между последовательно активирующимися центрами обработки.

Рис 4. Парные и симметричные относительно ретинотопической проекции участки коры головного мозга, распознающие лица и здания (категориальные модули)-А. Ретинотопические проекции стриарных и экстрастриарных отделов коры - Б. Схема внутримодульных и межполушарных взаимоотражающих связей обеспечивающих функционирование памяти и сознания.Временные модули обеспечивают передачу исходного сигнала для его сравнения с фиксированными в памяти шаблонами, в том числе для сохранения и анализа динамических образов.
Высказанные гипотезы только закладывают основы понимания алгоритмов сознания, разработка которых неизбежно приведет к созданию искусственного интеллекта. Однако, это будет непростой задачей. Смысловые сигналы в мозге, как шубой, окутаны активационным фоном, и их прямая регистрация крайне затруднительна.
В то же время, в отличие от сложного кодирования информационного потока, активирующие воздействия не несут смысловой нагрузки. Единственным их параметром является частота потенциалов действия (ПД). Чем выше частота, тем сильнее активация. Число активирующих элементов так же влияет на силу активации. Психостимуляторы и физические методы стимуляции (электростимуляция, магнитостимуляция и др.) влияют на два этих параметра. На этом основана терапия при дефиците активирующей системы. Психостимуляторы влияют на все отделы активирующей системы, что может вызвать нежелательные побочные эффекты при неправильном выборе дозы. Глубокая электростимуляция таламуса очень избирательна и эффективна при минимальных состояниях сознания [17].
Внешняя электрическая и магнитная стимуляция оказывает основное влияние на внутрикорковую активирующую систему вследствие особенностей анатомии мозга и физики распределения электрических и магнитных полей [4].
Таб. 1 Элементарные принципы эволюции и функционирования сознания.
· Принцип простого отражения(безусловный рефлекс)
· Принцип подкрепления(условный рефлекс)
· Принцип копирования (аффектор = эффектору)
· Принцип инверсии (+аффектор = -эффектору)
· Принцип ассоциаций и вариаций (наследование и полиморфизм)
· Принцип мультипликации и избыточности (множественные копии в пространстве и времени)
· Принцип спонтанности(свобода воли - следствие случайного выбора)
· Вероятностный принцип(странный аттрактор)
· Принцип симметрии(сложного множественного отражения)
· Принцип усиления (масштабирования)
· Сетевой принцип (адресация по содержимому)
· Синергетический принцип
· Модульная организация (инкапсуляция) -> Пространственное кодирование
· Принцип усложнения и динамической перестройки (простая система – простой ответ, сложная система – сложный многовариантный, многокомпонентный и более точный ответ)
· Формула сознания С = t(Ψ) ???
В отличие от сложностей, связанных с расшифровкой алгоритмов сознания (таб.1), понимание механизмов и значения активирующих воздействий уже сейчас может дать практический результат в лечении большого круга неврологических и психических расстройств.
Литература.
- Верхлютов В.М. Пространственная и временная вариабельность альфа-ритма магнитоэнцефалограммы у человека в норме. Журн.высш.нервн.деят. 1990. 40(5):944-949.
- Верхлютов В.М. , Щучкин Ю.В, Ушаков В.Л., Cтрелец В.Б., Пирогов Ю.А. Оценка локализации и дипольного момента источников альфа и тета – ритмов ЭЭГ с использованием кластерного анализа в норме и у больных шизофренией. Журн.высш.нервн.деят. 2006. 56 (1):47-55.
- Верхлютов В.М., Ушаков В.Л., Cтрелец В.Б. Фазовое опережение компонент ВП N170 у человека при повторном предъявлении изображений лиц. Журн.высш.нервн.деят. 2008 (В печати).
- Верхлютов В.М., Cтрелец В.Б. Попеременная активация височных долей мозга у человека при загрузке оперативной памяти. Журн.высш.нервн.деят. 2008 (В печати).
- Ковальзон В.М. Раскрыта природа нарколепсии. Природа. 2005. 11:3-8.
- Лорис С. Глаза открыты, мозг дремлет. В мире науки. 2007. (9):48-53.
- Ставцев A.Ю,.Ушаков В.Л.,.Верхлютов В.М, Моделирование влияния толщины слоев и проводимости тканей головы на потенциалы ЭЭГ с использованием метода конечных элементов. Журн.высш.нервн.деят., 2007, Т.57, N 6, с. 738-748.
- Филдз Р.Д. Вещественность белого вещества. В мире науки. 2008. (6):39-45. (R. Douglas Fields).
- Andersen P., Andersson S.A. Physiological Basis of the Alpha Rhythm. - Appleton-Century-Crofts, NY, 1968.
- DempseyE.W., Morison R.S. The production of rhythmically recurrent cortical potentials after localized thalamic stimulation. Am. J. Physiol. 1942. 135:293-300.
- Goldman R.I. et al., (2002) We found that increased alpha power was correlated with decreased MRI signal in multiple regions of occipital, superior temporal, inferior frontal, and cingulate cortex, and with increased signal in the thalamus and insula (фМРТ+ЭЭГ) .
- Levy I., Hasson U., Avidan G., Hendler T., Malach R. Centr-periphery organization object areas. Nature neuroscience. 2001. 4(5):533-.539.
- Mnatsakanian E.V., Tarkka I.M. Familiar-face recognition and comparison: source analysis of scalp-recorded event-related potentials. Clin.Neurophysiol. 2004. 115:880–886.
- Moruzzi G., Magoun H.W. Brain stem reticular formation and activation of the EEG. Electroencephalogr. Clin. Neurophysiol. 1949. 1: 455-473.
- Pfurtscheller G., Aranibar A. Event related cortical desynchronization detected by power measurements of scalp EEG. Electroencephalogr. Clin. Neurophysiol. 1977. 42:817-826.
16. Pfurtscheller, G. Event-related synchronization (ERS): an electrophysiological correlate of cortical areas at rest. 1992. Electroencephalogr. Clin. Neurophysiol. 83:62-69.
17. Schiff N.D. et al., fMRI reveals large-scale network activation in minimally conscious patient. Neurology. 2005. 64:514-523.