ринципы классификации многоклеточных
Главные направления биологической эволюции (ароморфоз, идиоадаптация, общая дегенерация). Биологический прогресс и пути его достижения. Биологический регресс (А.П.Северцов). Систематические группы. Естественная система органического мира как отражение эволюции. Основные закономерности эволюционного процесса: дивергенция, конвергенция, параллелизм, необратимость эволюционных преобразований. Результаты эволюции: многообразие видов, органическая целесообразность, постепенное усложнение организации.
Ученые полагают, что число вымерших видов растений и животных, когда-либо живших на Земле, в 5 - 9 раз больше числа современных видов. Такое многообразие ныне живущих и вымерших видов указывает на их значение в образовании высших систематических групп. ПРОЦЕСС ОБРАЗОВАНИЯ ИЗ НОВЫХ ВИДОВ НОВЫХ РОДОВ, ИЗ РОДОВ - НОВЫХ СЕМЕЙСТВ И Т.Д. НАЗЫВАЕТСЯ МАКРОЭВОЛЮЦИЕЙ. МАКРОЭВОЛЮЦИЯ СОВЕРШАЕТСЯ ЗА ИСТОРИЧЕСКИ ГРОМАДНЫЕ СРОКИ И НЕПОСРЕДСТВЕННОМУ НАБЛЮДЕНИЮ НЕДОСТУПНА. Однако принципиальных различий между микро- и макроэволюцией нет: в макроэволюции действуют те же самые факторы, что и в микроэволюции (видообразование).
В целом эволюционный процесс непрерывно идет в направлении наибольшего приспособления живых организмов к конкретным условиям среды. Смена условий приводит к замене одних приспособлений на другие. Это относится и к приспособлениям широкого характера. Например, появление легочного дыхания, двух кругов кровообращения, теплокровности, живорождения, млекопитания ...
Такие приспособления ведут к общему подъему организации, т.е. к биологическому прогрессу. ПРОГРЕСС ВСЕГДА СОПРОВОЖДАЕТСЯ УВЕЛИЧЕНИЕМ ЧИСЛЕННОСТИ, РАСШИРЕНИЕМ АРЕАЛА, ОБРАЗОВАНИЕМ НОВЫХ ПОПУЛЯЦИЙ, ПОДВИДОВ И ВИДОВ. БИОЛОГИЧЕСКИЙ РЕГРЕСС ОЗНАЧАЕТ СНИЖЕНИЕ УРОВНЯ ОРГАНИЗАЦИИ ЗА СЧЕТ УЗКОЙ СПЕЦИАЛИЗАЦИИ В ПРИСПОСОБЛЕННОСТИ ОРГАНИЗМОВ.В этом случае организмы теряют способность реагировать на изменения в окружающей среде,РЕГРЕСС ВЕДЕТ К УМЕНЬШЕНИЮ ЧИСЛА ОСОБЕЙ, ПОПУЛЯЦИЙ, ПОДВИДОВ, ВИДОВ, СУЖЕНИЮ АРЕАЛА. Чаще всего регресс ведет к вымиранию.
Советскими учеными А.Н. Северцевым (1866-1936) и И.И.Шмальгаузеном (1884-1963) был разработан вопрос о путях достижения биологического прогресса и выделено три основных направления. Это АРОМОРФОЗЫ - морфофизиологические изменения общего назначения, которые в процессе эволюции ведут к общему повышению уровня организации, но не являются узко приспособительными изменениями.
Ароморфозы дают возможность освоения совершенно новой среды обитания (например, выход на сушу из воды осуществился благодаря появлению легочного дыхания); обеспечивают большие преимущества в борьбе за существование (например, появление фотосинтеза, многоклеточности, полового процесса на ранних этапах развития органического мира).
В результате ароморфозов возникли крупные систематические группы - типы, классы, отряды.
Одновременно в ходе эволюции среди отдельных групп возникает большое количество мелких приспособлений к определенным условиям среды -ИДИОАДАПТАЦИИ. Например, приспособление одних цветков к опылению ветром, других к опылению насекомыми, третьих к самоопылению - все это проявление идиоадаптации. Путем идиоадаптаций образуются более мелкие систематические группы - семейства, роды, виды.
В процессе эволюции идиоадаптации возникают значительно чаще, чем ароморфозы.
Приспособительные изменения в процессе эволюции могут происходить путем снижения уровня общей организации. Резкое упрощение организации живого называется ОБЩЕЙ ДЕГЕНЕРАЦИЕЙ. Чаще всего дегенерация наблюдается при переходе к паразитическому или сидячему образу жизни и связана с тем, что окружающая Среда достаточно стабильна. Примерами могут служить известные вам из курса зоологии паразитические черви.
Несмотря на значительное упрощение организации, общая дегенерация не исключает биологического прогресса, т.е. виды, идущие по этому пути, могут увеличивать свою численность, расширять ареал и т.д. Однако дегенерация часто ведет к биологическому регрессу. По учебнику посмотрите соотношение различных направлений эволюции.
Если эволюционисты видят свою задачу в объяснении разнообразия организмов и истории их развития, а также в разработке общей теории эволюции, тоЦЕЛЬ СИСТЕМАТИКИ заключается в СОЗДАНИИ ТАКОЙ СИСТЕМЫ КЛАССИФИКАЦИИ, КОТОРАЯ ОТРАЖАЛА БЫ ЭВОЛЮЦИОННУЮ ИСТОРИЮ ВИДОВ.
В современной систематике растений и животных приняты следующие систематические группы. Для растений: отдел; класс; порядок; семейство; род; вид. Для животных: тип; класс; отряд; семейство; род; вид.
В основу современной естественной классификации органического мира положены признаки, свидетельствующие о родстве видов как с ныне живущими, так и с вымершими.
Самая маленькая в системе единица классификации - вид, но и он распадается на подвиды и популяции.
Попытки классифицировать организмы известны с древности. В прошлом, например, мир делили на царства растений, животных и минералов. До сих пор мы говорим: "Что это? - Растения", но "Кто это - Животные". В дальнейшем представление о том, что живое делится на два царства устарело. Сегодня в биологии выделяют два надцарства: прокариоты (бактерии, архебактерии) и эукариоты (растения, грибы, животные) и отдельно выделяются вирусы (доклеточные формы жизни).
Для описания видов и их классификации используются совокупность наиболее существенных признаков: анатомическое строение, особенности размножения, сложность организации (доклеточные, клеточные, безъядерные ядерные, одноклеточные, многоклеточные), сравнивают эмбриональное развитие, специфику химического состава (особенно белков) и физиологии, изучают поведенческие реакции, анализируют историческое развитие по ископаемым остаткам, проверяют прошлое и настоящее распространение по планете. Это позволяет в большинстве случаев определить положение вида в естественной системе классификации. ЕСТЕСТВЕННАЯ СИСТЕМА ОТРАЖАЕТ СТЕПЕНЬ РОДСТВА МЕЖДУ ГРУППАМИ ОРГАНИЗМОВ, Т.Е. ОТРАЖАЕТ ХОД ЭВОЛЮЦИИ ОРГАНИЧЕСКОГО МИРА НА ЗЕМЛЕ.
К основным закономерностям эволюционного процесса относятся дивергенция, конвергенция и параллелизм.
Еще Дарвин сформулировал ПРИНЦИП РАСХОЖДЕНИЯ ПРИЗНАКОВ.
РАСХОЖДЕНИЕ ПРИЗНАКОВ У РОДСТВЕННЫХ ФОРМ НАЗЫВАЕТСЯ ДИВЕРГЕНЦИЕЙ. В процессе дивергенции от одной исходной популяции берет начало целый ряд форм, образуя как бы ветви одного дерева. В бесконечном ряду поколений одни ветви отмирают, другие развиваются. Формы, наиболее расходящиеся по своим требованиям к условиям среды, имеют больше шансов выжить в ходе естественного отбора, так как они меньше конкурируют между собой, чем родоначальные или промежуточные группы. В результате дивергенции образуются подвиды, а затем и виды.
В эволюции наблюдается и другое явление - КОНВЕРГЕНЦИЯ ПРОЦЕСС НЕЗАВИСИМОГО ПРИОБРЕТЕНИЯ НЕРОДСТВЕННЫМИ ГРУППАМИ ОРГАНИЗМОВ СХОДНЫХ ПРИЗНАКОВ. Например, форма тела у рыб и дельфинов. Это сходство обусловлено общей средой обитания (море) и образом жизни.
Помимо дивергенции и конвергенции, в природе имеет место ПАРАЛЛЕЛИЗМ, - ПРИ КОТОРОМ НАБЛЮДАЕТСЯ НЕЗАВИСИМОЕ ПРИОБРЕТЕНИЕ СХОДНЫХ ЧЕРТ РОДСТВЕННЫМИ ОРГАНИЗМАМИ. В данном случае речь идет об общности происхождения и приспособлениях к одинаковой среде обитания. Например, морской котик, морж, тюлень. Наблюдаемые различия сложились в начале процесса видообразования.
Каковы же общие черты эволюционного процесса?ПРЕЖДЕ ВСЕГО, ЭТО ВОЗНИКНОВЕНИЕ "ЦЕЛЕСООБРАЗНОСТИ" ОРГАНИЗМОВ, Т.Е. ИХ СООТВЕТСТВИЯ УСЛОВИЯМ ОБИТАНИЯ И ИХ СПОСОБНОСТЬ МЕНЯТЬСЯ ПО МЕРЕ ИЗМЕНЕНИЯ ЭТИХ УСЛОВИЙ. ВО-ВТОРЫХ, ЭТО ПРОЦЕСС ВИДООБРАЗОВАНИЯ. В-ТРЕТЬИХ, ПОСТОЯННОЕ УСЛОЖНЕНИЕ ЖИЗНИ ОТ ПРИМИТИВНЫХ ДОКЛЕТОЧНЫХ ФОРМ ДО ЧЕЛОВЕКА.
За всю историю существования жизни на Земле обитало около 500 млн. видов. Сегодня их насчитывается около 2 млн. Это показывает, что совершенство современных форм - результат эволюции, в процессе которой погибли сотни миллионов видов. Эволюционные преобразования необратимы, любое упрощение или отклонение в организации ведет в конечном счете, к регрессу и вымиранию.
В мире, находящемся под угрозой разрушения в результате деятельности человека, важно как можно быстрее понять механизмы взаимодействия видов между собой и их реакции на создаваемые нами изменения, ибо это единственный способ избежать необратимых экологических изменений.
Подводя итог можно сказать о ПРАВИЛАХ ЭВОЛЮЦИИ. Их три.
ПРАВИЛО НЕОБРАТИМОСТИ ЭВОЛЮЦИИ. Эволюция - процесс необратимый. Группа организмов не может вернуться к прежнему состоянию, уже осуществленному в ряду их предков. Так в эволюции позвоночных на каком-то этапе от примитивных амфибий возникли рептилии, то они никогда не дадут начала амфибиям. Вернувшись к обитанию в океане, рептилии (ихтиозавры) и млекопитающие (киты) не стали рыбами.
Даже обратная мутация, по конкретному признаку приводящая к повторному возникновению данного гена не приводит к повторению всего генотипа в целом.
ПРАВИЛО ПРОГРЕССИРУЮЩЕЙ СПЕЦИАЛИЗАЦИИ. Группа, вступившая на путь специализации, как правило, в дальнейшем будут идти по пути ВСЕ БОЛЕЕ ГЛУБОКОГО ПРИСПОСОБЛЕНИЯ К УЗКИМ УСЛОВИЯМ СУЩЕСТВОВАНИЯ. Если в процессе эволюции одна из групп позвоночных приобрела адаптации к полету, то на последующем этапе эволюции это направление сохраняется и усиливается. Организм определенного строения не может жить в любой среде. В выборе адаптивной зоны или ее части группа ограничена особенностями своего строения.
ПРАВИЛО ЧЕРЕДОВАНИЯ ГЛАВНЫХ НАПРАВЛЕНИЙ ЭВОЛЮЦИИ. При рассмотрении главных направлений в эволюции групп ароморфозов и идиоадаптаций подчеркивалось регулярное чередование этих типов развития в эволюции.
ТАКИМ ОБРАЗОМ, ЭВОЛЮЦИЯ ПРЕДСТАВЛЯЕТ СОБОЙ НЕПРЕРЫВНЫЙ ПРОЦЕСС ВОЗНИКНОВЕНИЯ И РАЗВИТИЯ НОВЫХ АДАПТАЦИЙ. ОДНИ ИЗ ВНОВЬ ВОЗНИКАЮЩИХ АДАПТАЦИЙ ОКАЗЫВАЮТСЯ ОЧЕНЬ ЧАСТНЫМИ, И ИХ ЗНАЧЕНИЕ НЕ ВЫХОДИТ ЗА РАМКИ УЗКИХ УСЛОВИЙ. ДРУГИЕ ДАЮТ ВОЗМОЖНОСТЬ ВЫХОДА ГРУППЫ В НОВУЮ СРЕДУ И НЕПРЕМЕННО ВЕДУТ К БОЛЕЕ БЫСТРОМУ ЭВОЛЮЦИОННОМУ РАЗВИТИЮ ГРУПП В НОВОМ НАПРАВЛЕНИИ, К БОЛЕЕ ВЫСОКОЙ ОРГАНИЗАЦИИ.
Вопросы и задания для самоконтроля
1. В чем состоит биологический прогресс и биологический регресс?
2. Каковы главные направления биологической эволюции (прогресса)
3. В чем заключается биологическое значение ароморфозов и идиоадаптаций?
4. Чем характеризуется дегенерация?
5. Чем занимается наука систематика?
6. Верно ли утверждение о том, что современная систематика является отражением эволюционного процесса?
7. Перечислите основные закономерности эволюционного процесса.
8. Поясните биологическую сущность дивергенции, конвергенции, параллелизма.
9. Расскажите о результатах эволюции.
Подцарство одноклеточные (Protozoa)
Общая характеристика. В подцаретво объединены животные, по размерам микроскопические, а по общим чертам строения в большинстве одноклеточные. Исключение составляют колониальные, у которых тело образуют несколько клеток. Так, у вольвокса (рис. 29) число клеток в теле исчисляется тысячами, а их диффе-репцнровка на вегетативные и воспроизводящие приближает таких колониальных животных к многоклеточным. Тело иротозоа образовано протоплазмой с одним или несколькими ядрами, оргапел-лами и органами временными или постоянными (движения, пищеварения, выделения). Размножаются как бесполым, так и половым путем. По образу жизни — свободпожпвущпе и паразиты. Обитают в водоемах, почве, различных жидких и влажных средах. Могут переносить высыхание благодаря выделению защитных оболочек и в таком ипцистироваппом состоянии временно сохраняться. Иротозоа именуют прос т е й ш и м и, допуская условную трактовку наименования типа (греч. protos. норный).
В прошлом при малой осведомленности об этих существах они казались простыми в сравнении с многоклеточными. В настоящее время называть иротозоа простейшими можно, лишь отдавая дань истории.
Выяснено, что иротозоа имеют сложнейшую микроскопическую (и ультрамикроскопическую) организацию, обеспечивающую нее жизненные отправления. Обоснование в 30-х годах XIX в. клеточной теории строения животных и растений породило другое вольное толкование наименования типа, как объединяющего животных одноклет о ч н ы х. Термин «одноклеточные» не охватывает всего разнообразия форм, но удачен напоминанием о предках многоклеточных животных.
Протозоа — целостные организмы подобно тому, как целостными являются животные многоклеточные. Одноклеточпоеть и строении их тела есть своеобразная форма организации.
В основании родословного древа животных располагаются гипотетические примитивные первоначально безъядерные формы, которые в дальнейшем эволюционировали в клеточную форму с характерным для клетки признаком — наличием ядра – важного регулятора жизненных отправлений и носителя наследственной информации. В процессе эволюции одноклеточные дали начало животным многоклеточным. Их пути разошлись в очень давние времена.
Рис. 29. Вольвокс:
А — Volvoxglobular— участок колонии с половыми клетками; Б — Volvaxaureus— колония в процессе бесполого размножения; / — макрогаметы; 2 — мнкрогаметы; 3 — дочерние особи
Протозоа эволюционировали при сохранении родоначальной одноклеточной формы тела и микроскопических его размеров. Функции их органоидов, жизненные отправления в основном аналогичны жизненным отправлениям у многоклеточных. Последнее определяет возможность широкого использования протозоа для исследования многих биологических проблем, касающихся всего животного мира. Они стали объектами изучения ультраструктуры живой материи и ее основных жизненных свойств, явления анабиоза (скрытой жизни) и потенциальных возможностей сохранения жизни при крайних параметрах физико-химических факторов среды, изменчивости, экологии, разработки проблемы эволюции форм полового процесса, биологического анализа форм бесполого и полового размножения и пр.
Кажутся безграничными тончайшие морфофизиологические приспособления у протозоа к обстоятельствам жизни в разных экологических условиях, при разном образе жизни и поведении. Аналогичное известно и в мире многоклеточных. Сравнение общего и особенного у протозоа и метазоа в строении и жизненных отправлениях позволяет классифицировать тех и других полцарствами в царстве животных.
Подцарство Protozoa подразделяется на ряд типов, число которых продолжает уточняться систематиками. В данном учебнике рассматриваются следующие типы: Sarconiustigoplwra, Sporozoa,Cnidosporiciia, Mlcrosporidia, Ciliala.
Широкое распространение протозоа, своеобразие организации и практическое значение вызывали к ним большой интерес и повлекли накопление обширного материала. Со временем утвердились отрасли протозоологии: сельскохозяйственная, почвенная, ветеринарная, медицинская и другие, разрабатывающие теоретические проблемы и практическое использование полезных видов и меры ведения борьбы с вредными. Обитатели толщи воды океанов: фораминиферы (с известковыми раковинами), радиолярии (с кремневым скелетом), кокколиты (из жгутиковых, с известковым панцирем) — по отмирании образуют на дне мощные отложения известковых и кремниевых пород, входящих в состав земной коры. Мел на 90—98 % состоит из мельчайших панцирей кокколитофорид. Эти жгутиковые в живом виде содержатся в 1 м3 до 800 тыс. особей в Тихом океане, до 3 млрд.— в Атлантическом (Шумейко, 1978). Глобигериновый ил занимает в Мировом океане площадь около 120 млн. км2, имея толщину (местами) в несколько сот метров. Ископаемые протозоа используются в установлении стратиграфии осадочных пород как руководящие формы при определении возраста отложений, что важно в разведке полезных ископаемых и для исторической геологии. Установлено участие протозоа в почвообразовании, их санитарная роль в водоемах — как потребителей вредных бактерий, как извлекающих из воды ряд вредных веществ, поступающих в водоем с водосборной площади и со сточными водами. Медицинская и ветеринарная паразитология изучают биологию паразитов человека, домашних животных и возделываемых растений, их распространение, взаимосвязь с хозяином и другие вопросы общей и частной паразитологии. Выяснена возможность использования протозоа для испытания действия фитонцидов. Получен положительный результат выращивания культуры инфузорий для кормления мальков рыб в садках и прудах.
Строение и жизненные отправления. Размеры тела одноклеточных в большинстве микроскопические, измеряемые в микронах и миллиметрах. Исключение составляют лишь немногие, например морские корненожки — фораминиферы. Они тоже малых размеров, но отдельные виды современных достигают 3 см, а поперечник тела некоторых ископаемых превышает 10 см. Наименьшие размеры имеют те, что паразитируют в теле самих протозоа и в клетках различных органов многоклеточных животных. Из свободноживущих малые размеры характерны для почвенных обитателей, жизнь которых проходит в тончайшей пленке воды, окружающей твердые частицы земли. Они значительно меньше особей тех же видов из водоемов.
Форма тела весьма разнообразна (рис. 30), как это обычно наблюдается у животных очень крупной и богатой видами систематической категории. Причина тому — широкое распространение видов и их жизнь в разных экологических условиях.

Рис. 30. Различные формы тела нротозоа:
/—-амеба; 2 —- диффлугия; 5 — эвглена (Е.
oxyuris); 4 -— ламбдия; 5-стентор; £ — грегарииа
У хороших пловцов тело удлиненное, веретеновидное, обтекаемое, а у «парящих» в толще воды — с причудливыми выростами, увеличивающими поверхность тела, что наряду с включениями в протоплазме (жировыми каплями, газовыми пузырьками) способствует пассивному плаванию. Обитающие на дне водоемов отличаются относительно большой массивностью. У многих тело заключено в раковину, которая у одних органической природы, у других, как у диффлугии, построена из песчинок, сцементированных выделениями тела.
Свои особенности в строении эктопаразитов и эндопаразитов. Так, у трипанозом — паразитов крови тело веретеновидное, по его краю простирается протоплазматический парус, способствующий плаванию в вязкой плазме крови. Приспособительные черты в строении других паразитов крови выражены по-иному. Кишечные лямблии имеют тело уплощенное, с присасывательной площадкой на нижней стороне. Присоска удерживает паразита в кишечнике. Грегарины же (тоже кишечные паразиты) закрепляются выростами переднего отдела тела (крючьями, щупальце-образными придатками). Тело их вытянуто и подразделено на 2—3 отдела. Некоторые из грегарин наружной расчлененностью напоминают в миниатюре плоских членистых червей. Прикрепительным аппаратом обладают и эктопаразиты.
Рассмотренные черты морфологии протозоа, живущих в той или иной сходной среде, можно считать типичными лишь относительно по причине многообразия приспособлений, обусловленных отличиями в биологии — активностью поведения, различием в способах добывания пищи и т. д.
Покровы. На наружную пограничную зону тела приходятся такие важные для существования организма функции, как восприятие всевозможных воздействий извне, защита от механических и химических агентов внешней среды, поглощение кислорода, а при осмотическом питании — также поглощение пищи, удаление продуктов обмена веществ. Выполнение всех этих функций нельзя считать как зависящими только от структуры наружного покрова протоплазмы. В избирательной проницаемости покровов, как правило, участвует все тело одноклеточного животного. Выполнение функций определяется жизнедеятельностью всего организма и являет собою не механический, а физиологический процесс.
Тело корненожек, ряда эндопаразитов и многих других протозоа покрыто тончайшей мембраной. Большинство жгутиковых, инфузорий и саркодовых имеют покровы в виде тонкой, обычно эластичной пелликулы. Благодаря присутствию в пелликуле эластичных фибрилл она придает животному характерную для него форму, вместе с тем допуская временное ее изменение, и не препятствует совершать изгибы тела при движении. У ряда инфузорий пелликула достигает значительной толщины и плотности, приобретая свойства кутикулы или настоящего панциря (если пропитана солями). Вообще покровы протозоа отличаются сложностью и разнообразием строения. Как мембраны, так и пелликула составляют с протоплазмой одно целое и у некоторых видов в течение жизни или в отдельные ее периоды переходят одна в другую.
Под мембраной и пелликулой нередко отчетливо виден в световой микроскоп слой гомогенной желатинизированной протоплазмы— эктоплазмы. Глубже ее в теле находится жидкая и обычно зернистая эндоплазма. Она содержит ядро или ядерный комплекс из двух и более ядер, а также различные структуры, обычные для клеток и специфические, свойственные протозоа как организмам. Опорные фибриллы — один из видов скелетных образований. У некоторых на поверхности тела плотные, нерастяжимые волоконца и пластинки. Такие покровы приобретают свойства наружного скелета, выполняющего также защитную роль. Немногие протозоа имеют внутренний скелет. Как наружный, так и внутренний скелеты служат опорой для волокон (мионем), обладающих сократимостью. По функции они подобны миофиб-риллам — сократимым волокнам мышечных клеток метазоа. Скелет разных одноклеточных отличается и по строению, и по химическому составу. У корненожек он в виде домика, раковины. В строении раковин можно обнаружить весьма сложную структуру, как, например, у пресноводной раковинной амебы арцеллы.
Твердая основа ее раковины — кремний — органической природы, а входящая в нее эктоплазма имеет вид сети из призматических альвеол соответственно призматической структуре всего скелета. Электронный микроскоп позволяет отметить еще одну тонкую важную деталь ультраструктуры раковины — наличие сети нор, связующих эндоплазму с внешней средой.
Протозоа, ведущие жизнь в прикрепленном состоянии, снабжены скелетом, открытым на переднем конце, через который тело может выступать наружу. Этот участок тела служит для улавливания и приема пищи. Скелет плавающих в толще воды отличается тонкостью раковины и легкостью.
Трйхоцисты — специфические образования у инфузорий, расположенные под пелликулой. Они придают эктоплазме вид тонкой исчерченности. При раздражении инфузории кислотами, щелочами содержимое трихоцист вытягивается в длинные нити, смоченные веществом, вероятно, защитного действия. В электронный микроскоп видно, что нить имеет несколько утолщенную основную часть и острый конец. Трйхоцисты обычны для инфузорий, но изредка встречаются у жгутиковых. Их рассматривают как средство защиты и нападения. Трйхоцисты встречаются в теле животных то равномерно расположенными по всей периферии, как у инфузорий парамеций (рис. 31), то сконцентрированными в окружении рта. У некоторых они собраны в батареи, размещенные на определенных участках тела. В соответствии со строением и функцией среди трихоцист различают: пек-тицисты и токсицисты — орудие нападения хищных инфузорий, мукоцисты — слизистые и др. Трйхоцисты служат также для закрепления на субстрате во время питания.
Восстановление стрекательного аппарата после использования происходит путем новообразования внутри тела, на что указывает встречаемость трихоцист в эндоплазме.
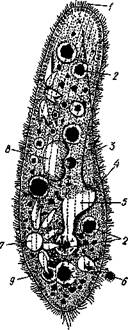
Рис. 31. Инфузория туфелька (Parameciumcau-datum):
/ — реснички; 2 — пищеварительные вакуоли; 3 — микронуклеус; 4 — ротовое отверстие; 5 — глотка; 5 —порошица; 7 — сократительная вакуоль (резервуар и радиальные приводящие каналы); 8—макронуклеус; 9 —трйхоцисты
Паразитические протозоа типа кни-доспоридий обладают стрекательным аппаратом, отличающимся большой сложностью и по строению напомииающим стрекательные капсулы низших многоклеточных животных (гидры и др.). Стрекательные капсулы ряда споровиков содержат спирально скрученную нить. У одних нить имеет трубчатое строение, у других устроена по-иному. При выстреливании один нити своим острым концом внедряются в тело жертвы, другие служат для заякоривания на теле или в кишечнике хозяина.
Аналогия в строении стрекательного аппарата споровиков и метазоа прослеживается и в том, что в образовании стрекательных капсул и у тех и других важное участие принимает аппарат Гольджи.
Раздражимость — восприятие факторов среды и реакция на их воздействие. Известна способность живой материи отвечать на разного рода воздействия характерным образом. Реакция живых существ на раздражение представляет собой не механический акт, а сложный биологический процесс. В живой природе ответная реакция на раздражение всегда специфична. На один и тот же фактор организм реагирует по-разному в зависимости от физиологического состояния в данный момент и от природных особенностей. Сытое одноклеточное животное, как и многоклеточное, иначе отвечает на пищевой раздражитель, чем голодное. Организмы, близко родственные, но живущие в разных средах, неодинаково реагируют на один и тот же раздражитель. Так, зеленые жгутиковые скапливаются в освещенной зоне водоема, а близкие к ним виды, но обитающие в затененных местах, лишенные хлорофилла, уходят от света.
Раздражимость у протозоа проявляется вне зависимости от высоты их организации, специфичности. Во всех случаях ответная реакция — функция всего организма.
Ответная реакция на раздражение движением называется таксисом, на свет — фототаксисом, на электричество — гальвано-таксисом (инфузории парамеции, сувойки и др. плывут в электрическом поле к катоду), на температуру — термотаксисом, на механический раздражитель — тигмотаксисом, на химический --хемотаксисом. Таксисы различают положительные (движение к раздражителю) и отрицательные (уход от раздражителя). Например, у амеб хемотаксис отрицательный к слабым щелочам, положительный термотаксис к оптимальной температуре.
Среди протозоа отличаются сложным поведением инфузории, имеющие и наиболее сложное строение тела. У ряда видов только в связи с питанием отмечено несколько форм движения. Сталкиваясь с резко неблагоприятными условиями, они могут двигаться задним концом и уходить от опасности.
Восприятие раздражений извне, рецепторная функция, осуществляется покровами и их производными: ресничками, жгутиками и пр., а также специальными органоидами, имеющимися у ряда видов. Так, органы зрения имеются у жгутиковых. Глазок (стигма) образован скоплением оранжево-красных капель, за которым располагается слой особо дифференцированной протоплазмы, а перед пигментным экраном — преломляющее свет тельце. В схеме такой глазок близок к схеме строения глаз у метазоа. У инфузорий описаны особые пузырьки с кристалликом внутри. Их называют статоцистами, а по функции принимают за органы равновесия. С изменением положения тела находящийся в пузырьке кристаллик, перемещаясь, давит на стенку пузыря в ином месте, что вызывает ответную реакцию восстановления положения тела в пространстве. Подобное же можно отметить в схеме строения органа равновесия многоклеточных. Находки статоцистов у инфузорий почти единичны, и оценка их функции требует дополнительных исследований.
Изучение поведения одноклеточных животных показало его сложность, неодинаковость по отношению к воздействующим факторам, привыканию к раздражителю (световому и др.).
Движение. Функцию движения у протозоа выполняют временные образования — псевдоподии и постоянные — жгутики и реснички.
Решающее значение иногда приобретают изгибы тела, как это свойственно ряду жгутиковых и др. Многие жгутиковые и инфузории из плавающих в толще воды поступательное движение совершают преимущественно за счет винтообразного вращения тела, направляемого движением жгутика или ресничек. В рамках каждого из отмеченных способов движения имеется значительное разнообразие с большим числом вариаций.
Движение у одноклеточных животных, как и у многоклеточных, связано с наличием особых контрактильных (сократительных) белков, отличающихся крупными молекулами волокнистой структуры. Такие белки (актиновые и мизиновые), ранее выявленные в миофибриллах мышечных клеток, обнаружены и в протоплазме протозоа, и в их органоидах движения — жгутиках и ресничках. Таким образом, в природе двигательной функции у животных того и другого подцарства прослеживаются аналогичные черты.
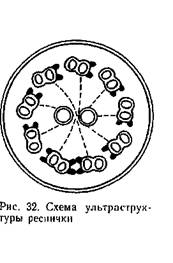
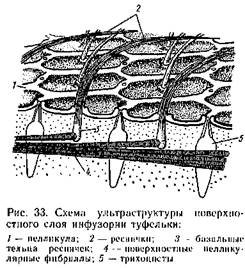

Рис. 34. Стилопихня
Ультраструктура жгутиков и ресничек протозоа, как и жгутиков и ресничек метазоа (в мерцательном эпителии, у сперматозоидов и пр.), показывает сходные черты в фибриллярном строении — наличие в каждой ресничке и в каждом жгутике 11 волоконец (нитей), расположенных продольно. Из них девять находятся на периферии и два занимают центральное положение (рис. 32).
Во всех случаях основание жгутиков и ресничек в пелликуле имеет вид гранулы, называемой базальным тельцем. У жгутиковых базальное тельце часто называют блефаропластом, а у инфузорий — кинетосомой. С базальным тельцем связаны особые фибриллы — корневые нити, которые у жгутиковых простираются в глубь тела, иногда достигая ядра.
Двигательный аппарат ряда высших протозоа (инфузорий) показывает сложность расположения фибрилл (рис. 33).
. Реснички отличаются от жгутиков многочисленностью иа теле и малой длиной. Усиление функций ресничек связано со слиянием их, что приводит к образованию толстых протоплазматических придатков наподобие ног (как у бегающих стилонихий, рис. 34), щупалец, мембран специального назначения, например в пред-ротовом углублении и т. д. Иногда подобное наблюдается и у жгутиковых.
Питание, добыча пищи, пищеварение. В тине протозоа встречаются всевозможные способы питания. Широко представлен древнейший способ — поглощение растворенных в воде веществ поверхностью тела. Такое питание осмотическим путем в мире животных называется сапрозойным. Оно распространено среди животных, обитающих в воде, богатой растворенными органическими и минеральными веществами, а также характерно для эндопаразитов. Столь же широко среди одноклеточных распространен собственно животный тип питания — голозойный, питание оформленной пищей, поглощение которой происходит с помощью специальных образований.

Рис. 35. Сосущие инфузории Didi-nium, напавшие на инфузорию
Растительный способ питания встречается преимущественно в классе жгутиковых у тех из них, которые имеют хроматофоры, содержащие пигменты: зеленый хлорофилл, желтый, синеватый и др. Их присутствие обеспечивает аутотрофам фотосинтез, продуктами которого обычно являются из углеводов крахмал и парамил. Ряд окрашенных жгутиковых при определенных обстоятельствах способны переходить к осмотическому и гетеротрофному питанию.
Материалы о зеленых жгутиковых свидетельствуют, с одной стороны, о наличии в подцарстве протозоа организмов, сочетающих в себе морфофизиологические черты двух царств органической природы: животных и растений, с другой — показывают своеобразие их организации и жизненных отправлений.
Органами добычи пищи и транспортировки ее внутрь тела служат: у корненожек — псевдоподии, у жгутиковых — жгутики и иногда близ его основания рот, у инфузорий — ресничный аппарат в предротовой области и реснички всего тела. Это общая схема. Инфузории в добыче пищи используют стрекательные нити, а сосущие инфузории — сосущий аппарат (рис. 35).
Добыча пищи с помощью псевдоподий признается наиболее примитивной (древней). Способность образовывать псевдоподии как временное явление сохранили некоторые одноклеточные из других классов. Амебы и другие корненожки захватывают пищу псевдоподиями (рис. 36) по-разному. Так, амебы могут обтекать пищевой комочек, окружать его своими псевдоподиями, могут активно втягивать его внутрь.

Рис. 36. Захват амебами пищевых частиц:
1-4 - амеба, захватывающая пищевую частицу; 5—7 — амеба, захватывающая нитчатую водоросль посредством «втягивания» ее внутрь тела
В пищевой рацион амеб входят водоросли иногда больших размеров, чем амеба. Электронный микроскоп позволил выяснить детали поглощения крупной добычи. Амеба, накрыв длинную водоросль своим телом, образует псевдоподии, так что водоросль оказывается в образованном ими канале. В стенках канала появляются микропсевдоподии, назначение которых — удерживать пищевой объект. Одновременно происходит втягивание нитчатой водоросли с закручиванием ее внутри тела амебы. Предполагается, что этому способствуют как сокращение самой протоплазмы, так и находящиеся в ней сократимые волоконца.
Прикрепленные к субстрату протозоа питаются преимущественно подвижной пищей, а плавающие — взвешенными в воде частичками детрита и живыми организмами. Скрепленные с субстратом сосущие инфузории способны улавливать своими щупальцами несколько проплывающих мимо инфузорий и поедать их (высасывать). Но бывает и так, когда плавающие хищные инфузории группой нападают на одну жертву и сообща высасывают ее. Среди протозоа есть монофаги и полифаги. Подтверждены экспериментально наличие у инфузорий выбора пищи и способность отличать на основе хеморецепции пищевые частицы — бактерий, дрожжей — от непищевых. В пищевой рацион некоторых протозоа входят и микроскопические животные из многоклеточных.
К дополнительным способам питания можно отнести поглощение продуктов фотосинтеза, продуцируемых симбиотическими хлореллами. Сожительство с ними широко представлено у протозоа. Их присутствие в теле одноклеточных сопровождается значительным повышением положительного фототаксиса (при условии оптимального освещения).
Пища, поступившая внутрь тела, обычно переваривается в пищеварительных вакуолях, которые возникают вместе с приемом пищи. Лишь у некоторых инфузорий (из панцирных) имеется внутри полость, внешне несколько напоминающая пищеварительный тракт метазоа. В случае осмотического питания вакуоли не образуются. Пищеварительные вакуоли содержат ферменты, воздействующие на белки (протеазы), на углеводы (карбогидра-зы) и пр., по функции аналогичные пищеварительным ферментам метазоа. Наблюдения за инфузориями и опыты с применением микрохимических реакций позволили установить железистый характер участков протоплазмы, прилегающих к вакуолям. В общем процесс пищеварения у протозоа аналогичен в самых основных чертах известному для метазоа, но многое остается невыясненным.
Длительность переваривания зависит от особенностей пищи и от ее размеров. Амеба, например, переваривает крупную нитчатую водоросль за 3—5 дней, а мелкую пищу — за несколько часов. Инфузория туфелька, питающаяся в основном бактериями, переваривает содержимое вакуолей примерно за час. Непереваренные остатки у той же инфузории накапливаются в определенном месте перед порошицей. При дефекации одновременно выбрасываются скопившиеся остатки из 4—5 вакуолей. У корненожек при отсутствии пелликулы и вообще более простой организации тела дефекация происходит в разных его участках.
Дыхание обеспечивает получение энергии путем поглощения кислорода для окисления органических веществ: углеводов, жиров и белков. Дыхание совершается с участием дыхательных ферментов (оксидазы, цитохрома и др.), т. е. подобно тому, как у многоклеточных. При распаде белков конечными продуктами будут С02, вода, азотистые вещества, в частности мочевина и мочевая кислота. Процесс дыхания (диссимиляция) в сочетании с ассимиляцией — основное свойство жизнедеятельности всякой живой материи. У некоторых диссимиляция осуществляется брожением, при котором происходит расщепление органических веществ с освобождением энергии.
Протозоа имеют неодинаковую потребность в кислороде. У инфузорий с их сложной организацией и поведением интенсивность газообмена значительно выше, чем у амеб. Кроме того, данные газообмена показывают приспособленность отдельных видов животных к существованию в условиях разной концентрации кислорода. Многие протозоа способны жить при ничтожном содержании его в среде. К таким относятся донные обитатели илистых грунтов в озерах и прудах, где и в летний сезон недостаток кислорода — обычное явление. К категории невзыскательных к кислороду животных надо причислить ряд паразитов, которые подобно дизентерийной амебе живут в кишечнике хозяина почти в анаэробных условиях. Примечательна способность протозоа водоемов временно переносить анаэробные условия в активном состоянии или впадая в анабиоз.
Выделение, или экскреция, служит освобождению организма от продуктов обмена веществ и избытка воды. Это происходит через поверхность тела и с помощью специальных периодически образующихся выделительных органов, которые у протозоа называются сократительными вакуолями или пульсирующими по причине ритмичного наполнения их с последующим выбрасыванием жидкости наружу. Вакуолей бывает одна или несколько. Сложность строения выделительной системы неодинакова. Амебы имеют только вакуоль, возникающую то тут, то там, а инфузории — вакуоль (или вакуоли) с системой приводящих каналов, как бы дренирующих участки тела во всех направлениях. Вакуоли занимают в инфузориях определенное место, и их опорожнение происходит также в определенном участке. Микрохимическими реакциями установлен железистый характер протоплазмы, прилегающей к приводящим каналам, свидетельствуя об экскреторном процессе выделения воды и продуктов распада. (Экскрецией называют выработку веществ, подлежащих удалению из тела, в отличие от секреции — процесса продуцирования веществ, нужных для жизнедеятельности организма.)
Сократительные вакуоли протозоа выполняют ряд функций. С их помощью выводится из организма избыток воды и тем поддерживается осмотическое давление в протоплазме на определенном уровне. Эта функция характеризует выделительную систему как осморегуляторпую. Жизнь пресноводных животных проходит в гипотонической среде, п аппарат осморегуляции для них необходим. Протоплазма содержит большее количество солей, чем пресная вода, п в силу разницы осмотического давления происходит непрерывное поступление воды внутрь тела через полупроницаемую оболочку покровов. Морские протозоа, ряд эндопаразитов и пр. из обитающих в изотопической среде сократительных вакуолей обычно не имеют. Наличие или отсутствие вакуолей определяется также регуляторпыми особенностями самих покровов. Частота пульсация значительно колеблется. Так, у амебы интервалы пульсации равны 1—5 мин, а у инфузорий — 10 20 с. Кроме осморегуляции, на экскреторные вакуоли возлагается удаление продуктом обмена веществ — углекислоты, мочевины и пр. В том случае, когда снята функция вывода воды, удаление продуктов диссимиляции и белкового обмена происходит через покровы. О значении сократительных вакуолей как осмо-регуляторов показывают опыты культивирования пресноводных протозоа в соленой воде. Постепенное наращивание солености влечет утрату амебами сократительных вакуолей, они их перестают образовывать.
Размножение протозоа происходит по-разному, и в особенностях его прослеживаются приспособительные черты. Нарастание числа особей у одноклеточных организмов происходит путем бесполого размножения (рис. 37). Наиболее обычной для евободпоживущих будет та его форма, при которой материнская особь разделяется па две дочерние (рис. 37, /). Ядро при этом делится кариокнпетпчеекнм путем.
Многие споровики и ряд паразитических протозоа других классов совершают бесполое размножение с той особенностью, что при делении материнской особи образуется несколько дочерних (множественное деление) (см. рис. 39). Высокая плодовитость — приспособительная черта биологии паразитов в связи с гибелью особей па пути распространения. При почковании после деления ядра следует неравномерное разделение протоплазмы и дочерний организм имеет вид почки па материнском теле (рис. 37, //). Среди инфузорий есть виды с внутренним почкованием. При этом дочерние особи образуются внутри камеры в теле матери, что напоминает живородпость (рис. 37, ///). Половое размножение метазоа происходит путем слияния двух половых клеток-гамет, а у протозоа гаметами становится особи, проходя этап созревания. У одних копулируют партнеры одинакового размера (изогамия), у других неодинакового (анизогамия) (рис. 38). У таких, как кокцидии, плазмодии, одни гаметы подобны яйцам, крупные, неподвижные, другие, как сперматозоиды метазоа, мелкие, подвижные. У вольвоксов подобно метазоа только часть клеток тела дают гаметы: мужские и женские, и есть виды раздельнополые п гермафродиты.
Своеобразен процесс педогамии и аутогамии. При педогамии копулируют дочерние особи, претерпев созревание. При аутогамии копулируют ядра одной и той же особи. После разделения ядра каждая часть проходит созревание (редукционное деление).
Половой процесс инфузорий — конъюгация. При конъюгации происходит временное сближение двух особей с взаимным оплодотворением ядрами, которые выполняют функцию гамет. Ядра в теле инфузорий двух категорий: одно крупное — макронуклеус, другое малое—микронуклеус. Первое называют вегетативным, второе — половым. В ходе конъюгации макронуклеус распадается, а микронуклеус проходит сложный процесс созревания, в результате которого в каждом из конъюгантов после двукратного деления микронуклеуса получаются четыре ядра, из которых три разрушаются, а четвертое делится, образуя два ядра (разных по величине). Более крупное остается на месте, а меньшее мигрирует в партнера и оплодотворяет оставшееся там ядро. Этим взаимным оплодотворением завершается конъюгация, и инфузории расходятся. Вскоре в каждой восстанавливается ядерный аппарат (рис. 39).



Среди инфузорий есть примеры, показывающие возможное сближение процессов копуляции и конъюгации, когда один из партнеров при конъюгации уменьшается в размерах и поглощается другим. В размножении ряда прикрепленных колониальных инфузорий видны приспособительные черты, выраженные в образовании на колонии особых веточек с конъюгантами. Нечто подобное известно у колониальных гидроидов в типе кишечнополостных.
Инцистирование — выделение на поверхности тела защитной оболочки — цисты. Заключение тела в цисту происходит по разным причинам: обычно с наступлением ухудшения условий жизни, иногда перед половым процессом или после приема пищи. У эндопаразитов инцистирование происходит обязательно перед выходом из хозяина во внешнюю среду. Оно способствует не только выживанию, но и распространению в пространстве (ветром, с помощью животных).
Онтогенез (греч. ontos— сущее, genesis— происхождение) — индивидуальное развитие от рождения до смерти. Естественным завершением жизни многоклеточного животного является смерть. Такая форма окончания жизни возникла в эволюции животного мира вместе с дифференцировкой тела на соматическую и воспроизводящую части. Для протозоа при их одноклеточном теле жизнь особи длится до разделения материнского организма на дочерние особи, представляющие собой новое поколение, которое растет и развивается до готовности к размножению.
У протозоа, имеющих в размножении стадию полового процесса, жизненный цикл совершается с чередованием поколений. Вслед за несколькими генерациями в результате бесполого размножения наступает появление половозрелого поколения. Так, инфузории парамеции производят до 60 поколений путем деления, прежде чем появится поколение конъюгантов. Бесполое размножение влечет возрастание числа особей и тем самым способствует сохранению вида. Развитие с чередованием поколений встречается и у метазоа, например у кишечнополостных.
К многоклеточным животным относятся все типы животных, кроме типа простейших. Различия между самыми простыми и наиболее сложными многоклеточными животными очень велики. Однако для всех них характерно разделение функций между различными группами клеток. В течение эволюции дифференциация тела животных на различные группы клеток, т. е. ткани, а затем на органы, состоящие из разных тканей, возрастала. Благодаря этому совершенствовалось (согласно закону единства и функции) выполнение функций, усиливалась (согласно закону корреляции) связь между разными частями тела, обеспечивалось лучшее приспособление организмов как целостных систем к неорганическим и органическим условиям среды обитания.
Многоклеточные животные, несомненно, произошли от простейших животных. Это заключение подтверждается следующими соображениями: 1) простейшие как более простые организмы возникли раньше многоклеточных животных и некоторые из них могли стать предками последних; 2) строение клеток и процессы их преобразования при бесполом и половом размножении у простейших и многоклеточных животных в основном очень сходны, что свидетельствует о родстве обоих подразделений царства животных; 3) первые стадии эмбрионального развития многоклеточных животных, как будет показано дальше, сходны по строению с некоторыми одноклеточными и колониальными простейшими, что свидетельствует (согласно биогенетическому закону) о происхождении самых низших многоклеточных животных от предков из типа простейших.
К какому же классу типа Protozoa принадлежали предки Metazoa? Казалось бы, это были инфузории, самые сложные из простейших с очень активным образом жизни. Однако для инфузорий характерен диморфизм ядер, который никогда не встречается в клетках многоклеточных животных. Кроме того, усложнение строения инфузорий происходило в пределах одной клетки, что затрудняло бы дифференциацию подобных клеток в разных направлениях. Споровики не могли дать начало более высокой группе животных, так как паразитический образ жизни никогда не способствовал общему подъему организации. Ложноножковые — медлительные простейшие, для них характерно образование раковин, защищающих их от хищников, а не активизация образа жизни. Поэтому, по мнению большинства ученых, многоклеточные животные произошли от весьма активных гетеротрофных жгутиковых.
Гипотез о происхождении Metazoa было предложено много, из которых наибольшее распространение получили две: известного немецкого зоолога Э. Геккеля (1834—1919) и выдающегося русского биолога И. И. Мечникова (1845—1916). В основе этих гипотез лежит использование биогенетического закона, так как организмы, промежуточные между жгутиковыми и самыми простыми многоклеточными животными, не имели твердых частей, могущих сохраниться в ископаемом состоянии, и судить о их строении можно главным образом путем изучения начальных стадий эмбрионального развития многоклеточных животных. Поэтому необходимо вкратце напомнить об этих стадиях.
Развитие многоклеточных животных начинается с зиготы, которая делится на 2, 4, 8, 16 и большее количество клеток, или бластомеров, остающихся соединенными. В результате описанных процессов деления бластомеров образуется кучка клеток, часто называемая морулой. Деление бластомеров продолжается и возникает шарообразная (или другой формы) бластула с полостью (бластоцелем) внутри, окруженной одним слоем клеток. Стадии развития от зиготы до бластулы включительно принято называть дроблением зиготы. В 70-х годах прошлого столетия, когда Геккель разрабатывал свою гипотезу на основании исследований эмбрионального развития-разных групп животных, было установлено, что бластула вследствие впячивания (инвагинации) части клеток в бластоцель и прилегания их к оставшимся снаружи клеткам превращается в зародыши с двухслойными стенками, окружающими полость с отверстием наружу. По окончании эмбрионального развития упомянутая полость превращается в кишечную, а отверстие ее становится ртом. Эта стадия получила название гаструлы. Наружный слой ее принято называть эктодермой, внутренний, окружающий кишечную полость,— энтодермой. На основании изложенных данных Геккель предположил, что предками многоклеточных животных были одноклеточные простейшие, затем колонии в виде кучек клеток, похожие на морулу, далее шарообразные колонии, сходные с бластулами и названные блас-теями, и наконец самые простые многоклеточные животные, похожие на гаструл и названные гастреями. Однако Геккель не учел, что эмбриологические данные других ученых (сам он эмбриологом не был), которыми он пользовался для обоснования своей гипотезы, относились к зародышевому развитию уже более сложных многоклеточных животных, и в начальных стадиях их онтогенеза могли произойти серьезные изменения (вплоть до выпадения какой-нибудь стадии, как это иногда наблюдается в связи с необходимостью сокращения эмбрионального развития). Кроме того, согласно гипотезе Геккеля, гастреи были уже не примитивными многоклеточными животными, а имели кишечную полость, т. е. резко отличались от бластей. Очевидно, между бластеями и гастреями должна была быть стадия более примитивных многоклеточных животных, чем гастреи.
И. И. Мечников, который в течение многих лет занимался исследованиями эмбрионального развития различных групп многоклеточных животных, вскоре после опубликования гипотезы Геккеля обратил особое внимание на изучение зародышевого развития низших современных типов Metazoa— губок, кишечнополостных и примитивных плоских червей. Он доказал, что у этих животных отдельные клетки развившейся бластулы проникают внутрь бластоцеля и заполняют его. В результате образуется зародыш, состоящий из двух разных слоев — наружного и внутреннего, не имеющий ни рта, ни кишечной полости. Мечников назвал эту стадию паренхимулой и предложил следующую гипотезу происхождения первых многоклеточных животных.
Властей были планктонными колониями простейших, состоящих из одинаковых с тонкими оболочками клеток, которые могли выпускать ложноножки, превращающиеся в простые жгуты, и выполняли все функции (движение, захват мелких организмов и органических кусочков, переваривание их и т. д.). Соединения этих клеток были рыхлыми, между ними имелись поры, и клетки могли менять свое положение в колонии. Подобные передвижения клеток наблюдаются у низшего типа многоклеточных животных — губок и даже у представителей более сложных типов. Поэтому отдельные клетки бластей, захватившие ложноножками мелкие организмы или органические кусочки, могли мигрировать в бласто-цель и там переваривать захваченную пищу, после чего возвращались на поверхность колоний. Другие клетки, захватившие пищу, погружались в бластоцель и т. д. При описанных миграциях клеток в бластеях намечалось разделение функций, но только временное: одни и те же клетки, будучи на поверхности колонии, выполняли одни функции, а внутри нее — другие. Такое разделение функций недостаточно. Клетки, обеспечивающие движение и защиту, должны иметь хорошо развитые жгуты и плотную оболочку, а клетки, захватывающие пищу,— тонкую оболочку, позволяющую выпускать ложноножки. Поэтому в результате длительного естественного отбора выжили те потомки бластей, у которых закрепилось более или менее постоянное разделение функций между клетками: одни клетки образовали наружный слой, у них развились более сложные жгутики и плотные оболочки; другие клетки, сохранившие тонкие оболочки и способность выпускать ложноножки, образовали внутренний слой. Ни рта, ни кишечной полости у этих первых многоклеточных животных еще не было. В наружном слое было много пор, а внутренний слой был очень рыхлый (между клетками имелись небольшие промежутки). Это позволяло им двигаться, выпускать ложноножки и захватывать мелкие организмы и органические кусочки, попадающие внутрь тела подобных организмов через поры.
И. И. Мечников назвал этих гипотетических (предполагаемых) первичных многоклеточных животных паренхимеллами, имея в виду их сходство с паренхимулами. Наружный слой паренхимелл он предложил назвать кинобластом (греч. «киное» — движение), а внутренний — фагоцитобластом (греч. «фагос» — пожирающий). Следовательно, предлагая такие термины, он отмечал главные физиологические функции обоих слоев паренхимелл. Рассматриваемая гипотеза была поддержана на основании разносторонних глубоких исследований эволюции тканей многоклеточных животных А. А. Заварзиным (1886—1945) и получила дальнейшее развитие в трудах А. В. Иванова и других советских и иностранных ученых. Особенно важно отметить, что в 70—80-е годы текущего столетия было доказано существование в настоящее время многоклеточных животных, в основном похожих на гипотетических паренхи-мелл, названных трихоплаксами. Это морские животные малых размеров (2—4 мм), имеющие форму тонких пластинок, окруженных одним слоем жгутикового эпителия. Вся внутренность этих животных заполнена амебовидными и другими клетками. Трихоплаксы, медленно меняя свою форму, ползают по поверхности водорослей и других субстратов. Сторона, обращенная к субстрату (брюшная), отличается по строению от противоположной стороны (спинной). Передний и задний концы тела не выражены.
Экспериментально доказано, что внутренние амебовидные клетки трихоплаксов, подходя к поверхностному слою, способны поглощать мелкие организмы (в опытах использовались дрожжи). Размножение бесполое (путем деления на два одинаковых организма или образованием многоклеточных «бродяжек», т. е. плавающих зародышей) и половое. От паренхимелл, какими их представлял себе Мечников, трихоплаксы отличаются переходом к ползающему образу жизни на разных донных субстратах. Вероятно, паренхимеллообразные животные были вытеснены из толщи воды произошедшими от них гастреями, у которых начали развиваться примитивные нервные и мышечные клетки. Благодаря этому они могли быстро захватывать пищу, которая попадала в кишечную полость. Последняя образовалась путем Постепенного раздвижения клеток фагоцитобласта и сообщалась с наружной средой только одним отверстием — ртом. Поры в стенках тела закрылись, что защищало гастрей от проникновения в их тело вредных организмов.
Согласно гипотезе И. И. Мечникова, исходным предком для всех многочисленных животных были паренхимеллы, тела которых состоят из клеток, способных к фагоцитозу. Руководствуясь этим положением, он предпринял поиски фагоцитарных клеток у разных многоклеточных животных и доказал, что они имеются у всех изученных им групп Metazoa. На основании этих исследований и ряда опытов он создал теорию фагоцитоза, согласно которой в защите животных от инфекционных заболеваний огромную роль играют особые клетки — фагоциты, которые, выпуская ложноножки, пожирают бактерии или способствуют их гибели, выделяя особые ядовитые вещества. Фагоциты выполняют также санитарную роль, уничтожая отмирающие клетки организма. Так, исследования выдающегося русского биолога-дарвиниста, предпринятые им для разрешения проблемы происхождения многоклеточных животных, привели к оформлению теории, имеющей столь большое значение для медицины и ветеринарии. В своих работах по фагоцитозу вместо термина паренхимелла И. И. Мечников предпочитал пользоваться термином фагоцителла.
ринципы классификации многоклеточных
Многоклеточных животных делят на три надраздела: фагоци-теллообразные с единственным типом пластинчатых, паразои с одним типом губок и эуметазои, или настоящие многоклеточные.
Фагоцителлообразные, включающие только два вида мелких плоских морских животных, представляют собой наиболее примитивных многоклеточных. Их тело не имеет определенных очертаний, как у амебы, у них нет ни переднего, ни заднего конца и очень слаба дифференциация клеток. Многие зоологи считают их близкими по организации к первым многоклеточным животным.
В отличие от фагоцителлообразных, у паразоев существует ясная дифференциация клеток, но нет настоящих тканей и органов.
Дифференцированные ткани и органы появляются у эуметазоев, которые, в свою очередь, делятся на два раздела — лучистых (кишечнополостные и гребневики) с радиальной симметрией тела и билатеральных — двусторонне-симметричных животных. В таблице 10 приведена сравнительная характеристика многоклеточных, не относящихся к билатеральным.
Классификация билатеральных животных базируется на понятии о полости тела, представляющей собой пространство между стенкой тела и кишечником. У плоских червей это пространство занято паренхимой и полости тела, по существу, нет. У других же билатеральных эта полость есть. В одном случае она заполнена жидкостью и лишена собственной эпителиальной выстилки. Таковы, например, круглые черви, особый тип, относящийся к первич-нополостным животным. В другом случае полость тела имеет собственные клеточные стенки, которые формируются в онтогенезе из третьего зародышевого листка, или мезодермы. Эту полость называют вторичной, или целомом. Все высшие являются целомическими, или вторичнополостными, животными.
Таблица 10.
Сравнительная характеристика трех низших типов многоклеточных
| Признаки | Надраздел Phagocytellozoa Тип Placozoa | Надраздел Parazoa Тип Porifera | Надраздел Eumetazoa Тип Coelenterata |
| Зародышевые листки | отсутствуют | оформление зародышевых листков; инверсия листков в онтогенезе | два зародышевых листка: эктодерма и энтодерма |
| Симметрия | асимметрия | осевая с гетеро-полярной осью и асимметрия | радиальная симметрия с гетеро-полярной осью |
| Типы вегетативных клеток | жгутиковые (кинобласт) и веретеновидные (фагоцитобласт) | пинакоциты, хоаноциты, амебоидные, склеробласты, спонгиобласты | эктодерма: эпителиально-мышечные, нервные, стрекательные; энтодерма: эпителиально-мышечные, железистые |
| Развитие личинки | без выраженного метаморфоза; молодые стадии — «бродяжки» | с метаморфозом; личинки: амфи-бластула, цело-бластула, паренхимула | с метаморфозом, реже прямое; личинка: планула |
Билатеральных животных также обычно делят на два подраздела — первичноротых и вторичноротых, резко отличающихся эмбриональным развитием. У первичноротых бластопор остается ртом и у взрослого животного. Мезодерма формируется из пары клеток — телобластов зародыша.
У вторичноротых на месте бластопора образуется анальное отверстие взрослого животного, а постоянный рот закладывается независимо от бластопора и часто на противоположной стороне. Мезодерма же образуется путем выпячивания боковых стенок кишечника (энтероцельно). Ко вторичноротым принадлежат типы иглокожих, полухордовых и хордовых. Остальные типы билатеральных относят к первичноротым.
Классификация многоклеточных
Надраздел Фагоцителлообразные — Phagocytellozoa
Тип Пластинчатые — Placozoa
Надраздел Паразои — Parazoa
Тип Губки — Porifera, или Spongia
Надраздел Эуметазои — Eumetazoa
Раздел Лучистые — Radialia
Тип Кишечнополостные — Coelenterata
Тип Гребневики — Ctenophora
Раздел Билатеральные — Bilateria
Подраздел Первичноротые — Protostomia
Тип Плоские черви — Plathelmintes
Тип Круглые черви — Nemathelminthes
Тип Головохоботные — Cephalorhyncha
Тип Коловратки — Rotifera
Тип Скребни — Acanthocephala
Тип Немертины — Nemertim
Тип Камптозои — Kamptozoa
Тип Кольчатые черви — Annelida
Тип Эхиуриды — Echiurida
Тип Сипункулиды — Sipunculida
Тип Членистоногие — Arthropoda
Тип Моллюски — Mollusca
Тип Онихофоры — Onichophora
Тип Язычковые, или Пятиустки — Linguatulida, или Pentastomida
Тип Тихоходки — Tardigrada
Тип Форониды — Phoronida
Тип Плеченогие — Brachiopoda
Тип Мшанки — Bryozoa
Тип Щетинкочелюстные — Chaetognatha
Тип Погонофоры — Pogonophora
Подраздел Вторичноротые ~ Deuterostomia
Тип Иглокожие — Echinodermata
Тип Полухордовые — Hemichordata
Тип Хордовые — Chordata
Следует сказать, что система типов беспозвоночных животных до сих пор окончательно не устоялась. Зоологи регулярно выделяют новые типы животных и среди уже известных групп, и в результате открытия новых животных, а также спорят о статусе прежних типов. В учебнике рассматриваются наиболее важные типы беспозвоночных.
Двусторонне-симметричные животные (билатеральные)
 Двусторонне-симметричные или билатеральные (лат. Bilateria) — раздел животных, к которому относятся все двусторонне-симметричные типы. У всех них левая сторона тела зеркальным образом соответствует правой. Этот принцип, однако, не относится к отдельным внутренним органам. Другое важное исключение составляют иглокожие, взрослые формы которых приближаются к радиальной симметрии, в то время как личинки билатерально симметричны.
Противоположенным к разделу двусторонне-симметричных является раздел кишечнополостных, которые построеннырадиально-симметрично, а также представители иных подцарств, чем эуметазои. Губки и пластинчатые не проявляют симметрию. Двусторонне-симметричные или билатеральные (лат. Bilateria) — раздел животных, к которому относятся все двусторонне-симметричные типы. У всех них левая сторона тела зеркальным образом соответствует правой. Этот принцип, однако, не относится к отдельным внутренним органам. Другое важное исключение составляют иглокожие, взрослые формы которых приближаются к радиальной симметрии, в то время как личинки билатерально симметричны.
Противоположенным к разделу двусторонне-симметричных является раздел кишечнополостных, которые построеннырадиально-симметрично, а также представители иных подцарств, чем эуметазои. Губки и пластинчатые не проявляют симметрию.
|
У самых примитивных билатеральных животных - низших червей (к которым относятся плоские и круглые черви) появляется третий зародышевый листок – мезодерма. Это крупный ароморфоз, благодаря которому появляются дифференцированные ткани и системы органов [наверное, он возник у единого предка билатеральных, развившегося из губкообразного прародителя].
Следующим крупным ароморфозом является вторичная полость тела, или целом. Благодаря этому становится возможным разделение тела на отделы
Затем эволюционное древо животных разветвляется на первичноротых и вторичноротых.
(На основе Википедии и других маиериалов)
Первичноротые животные
Первичноротые (лат. Protostomia) — подраздел двусторонне-симметричных животных. В период зародышевого развития на месте их первичного рта (бластопора) образуется рот или, при щелевидном замыкании бластопора, рот и анальное отверстие. Этим они отличаются от вторичноротых, у которых на месте бластопора образуется анальное отверстие, а ротовое возникает позже в другом месте.
У наиболее примитивных первичноротых животных (кольчатых червей и им подобных) появились примитивные конечности (параподии) и гомономная сегментация тела.
(Из Википедии и других источников)
Раньше плоские черви считались предками круглых червей, а кольчатые черви — предками членистоногих. По последним данным плоские черви оказались родственниками кольчатых, а круглые — членистоногих.
Анализ генов подтвердил теорию, согласно которой разделение на первичноротых и вторичноротых произошло еще до того, как у животных сформировался целом (вторичная полость тела). Первичноротые подразделяются на 2 четкие эволюционные линии:
- Lophotrochozoa ("спиральные" - плоские и кольчатые черви, моллюски, брахиоподы, немертины)
- и Ecdysozoa ("линяющие" - круглые и головохоботные черви, членистоногие, онихофоры, тихоходки).
Филогенетическое дерево вторичноротых животных
Вторичноротовые животные (дейтеростомы)
/ / \ \ \
/ / \ \ \
Щетинкочелюстные Полухордовые Иглокожие Погонофоры Хордовые
Происхождение вторичноротых животных
Происхождение вторичноротых неясно. Возможно они произошли от радиальных (кишечнополостных) животных независимо от первичноротых. По другим гипотезам, предками вторичноротых были представители одного из примитивных типов первичноротых, объединяемых в группу низших червей. Последние данные молекулярных исследований говорят в пользу первой теории.
У вторичноротых в период зародышевого развития на месте первичного рта (бластопора) образуется анальное отверстие, а собственно рот независимо появляется в передней части тела. Есть вторичная полость тела (целом).
(Из Вторичноротые в Википедии)