Рефлексы регулирующие мышечный тонус
Мышечная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы тела. Эти процессы регулируются различными отделами ЦНС.
Роль спинного мозга
Мышечный тонус является по своей природе рефлекторным актом. Для его возникновения достаточна рефлекторная деятельность спинного мозга. При длительном растяжении мышц в поле силы тяжести возникает постоянное раздражение их проприорецепторов, потоки импульсов от которых проходят по толстым афферентным волокнам в спинной мозг, где передаются непосредственно (безучастия вставочных нейронов) на альфа-мотонейроны передних рогов и вызывают тоническое напряжение мышц. Такие двухнейронные (или моносинаптические) рефлекторные дуги лежат в основе тонических сухожильных (с рецепторов сухожилий) и миотатических рефлексов на растяжение (с рецепторов мышечных веретен). Это рефлексы активного противодействия мышцы ее растяжению. В произвольной двигательной деятельности человека иногда требуется подавление этих рефлексов, например, при выполнении шпагата.
Степеньтонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней альфа-мотонейронами. Однако, потоки этих импульсов могут регулироваться вышележащими этажами нервной системы, в частности, неспецифическими отделами ствола мозга с помощью так называемой гамма — регуляции. Разряды гамма-мотонейронов спинного мозга под влиянием ретикулярной формации повышают чувствительность рецепторов мышечных веретен. В результате при той же длине мышцы увеличивается поток импульсов от рецепторов к альфа-мотонейронам и далее к мышце, повышая ее тонус.
Роль коры головного мозга, мозжечка и ствола мозга
В регуляции тонуса участвует также медленная часть пирамидной системы и различные структуры экстрапирамидной системы (подкорковые ядра, красные ядра и черная субстанция среднего мозга, мозжечок, ретикулярная формация ствола мозга, вестибулярные ядра продолговатого мозга).
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел ретикулярной формации среднего мозга, а угнетение — тормозящий отдел продолговатого мозга. В отличие от нее специфическая система (моторные центры коры больших полушарий и ствола мозга) действует избирательно, т. е. на отдельные группы мышц-сгибателей или разгибателей. Усиление тонуса мышц-сгибателей вызывают корковые влияния, передающиеся непосредственно к нейронам спинного мозга но корково-спинномозговой (пирамидной) системе, а также через красные ядра (по корково-красноядерно-спинномоз-говой системе) и частично через ретикулярную формацию (по ретикуло-спинномозговой системе). В противоположность им, влияния, передающиеся через вестибулярные ядра продолговатого мозга к вставочным и двигательным нейронам передних рогов спинного мозга (по вестибуло-спинномозговым путям), вызывают тоническое повышение возбудимости мотонейронов мышц-разгибатслей. что обеспечивает повышение тонуса этих мышц.
Мозжечок формирует правильное распределение тонуса скелетных мышц: через красные ядра среднего мозга он повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга — усиливает тонус мышц-разгибателей. В поддержании позы и равновесия тела, регуляции тонуса мышц основное значение имеет медиальная продольная зона мозжечка — кора червя. При мозжечковых расстройствах падает тоническое напряжение мышц (атония) и вследствие ненормального распределения тонуса мышц конечностей возникает нарушение походки (атаксия).
Бледное ядро угнетает тонус мышц, а полосатое тело снижаем его угнетающее действие.
Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы тела, обеспечивается ее соответствие двигательной задаче. Непосредственное отношение к регуляции тонуса мышц имеют медленные пирамидные нейроны положения. Корковые влияния на тонические реакции мышц передаются через медленную часть пирамидного тракта и через экстрапирамидную систему.
Рефлексы поддержания позы (установочные)
Специальная группа рефлексов способствует сохранению позы — это так называемыеустановочные рефлексы. К ним относятся статические и стато-кинетические рефлексы, в осуществлении которых большое значение имеют продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменениях положения головы в пространстве — лабиринтные рефлексы, возникающие при раздражении рецепторов вестибулярного аппарата. 2) шейные рефлексы — возникающие с проприорецепторов мышц шеи при изменении положения головы по отношению к туловищу, и 3) выпрямительные рефлексы — с рецепторов кожи, вестибулярного аппарата и сетчатки глаза. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус мышц-сгибателей (лабиринтный рефлекс). С помощью выпрямительного рефлекса происходят последовательные сокращения мышц шеи и туловища, а затем и конечностей. Этот рефлекс обеспечивает вертикальное положение тела теменем кверху. У человека он проявляется, например, при нырянии.
Стато-кинетические рефлексыкомпенсируют отклонения тела при ускорении ши замедлении прямолинейного движения (лифтный рефлекс), а также при вращениях(отклонения головы, тела и глаз в сторону, противоположную движению). Перемещение глаз со скоростью вращения тела, но в противоположную сторону, и быстрое их возвращение в исходное положение — нистагм глаз — обеспечивает сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
Роль различных отделов ЦНС в регуляции движений
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
Спинной мозг осуществляет ряд элементарных двигательных рефлексов:рефлексы на растяжение (миотатические и сухожильные рефлексы, например, коленный рефлекс), кожные сгибательные рефлексы (например, защитный рефлекс отдергивания конечности при уколах, ожогах), разгибательные рефлексы (рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты — регуляцию деятельности мышц-антагонистов, ритмических и шагательных рефлексов, лежащих в основе локомоций и других движений.
Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, по и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — торможение. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбуждаются центры разгибателей. Такие координационные взаимоотношения между спинальными моторными центрам и названы реципрокной (взаимосочетанной) иннервацией мышц-антагонистов. Однако реципторные отношения между центрами мышц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Составной частью различных сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Это одна из форм древних и относительно простых рефлексов. Они особенно выражены при выполнении циклической работы, включаются в шагательные рефлексы. Основные механизмы шагательных движений заложены в спинном мозге. Специальные нейроны (спинальпые локомоторные генераторы) и многочисленные взаимосвязи внутри спинного мозга обеспечивают последовательную активность различных мышц конечностей, согласование ритма и фаз движений, приспособление движений к нагрузке на мышцы. В среднем мозгу расположены нейроны «локомоторной области», которые включают этот механизм и регулируют мощность работы мышц, обеспечивая примитивную форму локомоций — без ориентации в пространстве.
Нейроны промежуточной продольной зоны коры мозжечка согласуют позные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации моторных программ к текущей ситуации. Программирование каждого последующего шага осуществляется ими на основе анализа предыдущего. Кроме того производится согласование движений рук и ног, и особенно — регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четкомподдержании темпа ритмических движений объясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и походящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации, влияющие на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных) в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы — с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений, четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
Роль различных отделов коры больших полушарий
Функцией комплекса различных корковых областей является определение целесообразности локомоций, их смысла, ориентации в пространстве, перестройка программ движений в различных ситуациях, включение ритмических движений как составного элемента в сложные акты поведения. Об участии различных корковых областей в регуляции циклических движений можно судить по появлению в их электрической активности медленных потенциалов в темпе движения — «меченых ритмов»ЭЭГ, а при редких движениях — по изменениям огибающей амплитуду ЭЭГ кривой.
В организации двигательных актов участвуют практически все отделы коры больших полушарий. Моторная область коры (прецентральная извилина) посыпает импульсы к отдельным мышцам, преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт («кинетическую мелодию»)осуществляютвторичные поля премоторной области. Они определяют последовательность двигательных актов, формируют ритмические серии движений, регулируют тонус мышц. Постцентральная извилина коры представляет собой общечувствительное поле, которое обеспечиваетсубъективное ощущение движений. Нижнетеменные области коры (задние третичные поля) формируют представления о взаимном расположении различных частей тела и положении тела в пространстве, обеспечивают точную адресацию моторных команд к отдельным мышцам и пространственную ориентацию движений. Области коры, относящиеся к л им бич ее кой системе (нижние и внутренние части коры), ответственны за эмоциональную окраску движенийи управление вегетативными их компонентами.
В высшей регуляции произвольных движений важнейшая роль принадлежитпередне-лобным областям (передним третичным полям). Здесь помимо обычных вертикальных колонок нейронов существует принципиально новый тип функциональной единицы — в форме замкнутого нейронного кольца. Циркуляция импульсов в этой замкнутой системе обеспечивает кратковременную память. Она сохраняет в коре возбуждение между временем прихода сенсорных сигналов и формированием ответной эфферентной команды. Такой механизм служит основой сенсомоторной интеграции при программировании движений, при осуществлении зрительно-двигательных реакций.
Функцией передне-лобной (третичной) области коры является сознательная оценка текущей ситуации и предвидение возможного будущего, выработка цели и задачи поведения, программирование произвольных движений, их контроль и коррекция. Соответствие выполняемых действий поставленным задачам придает движениям человека определенную целесообразность и осмысленность. При поражении лобных долей движения человека становятся бессмысленными.
Речевая регуляция движений
Спецификой регуляции движений у человека является то, что они подчиненыречевым воздействиям, т. е. могут программироваться лобными долями в ответ на поступающие извне словесные сигналы, а также благодаря участию внешней или внутренней речи (мышления) самого человека. В этой функции принимают участие расположенные в левом полушарии человека сенсорный центр речи Вернике и моторный центр речи — центр Брока. Считают, что афферентная импульсация от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на речевой основе избирательные связи в коре облегчают составление моторных программ.
Эта управляющая система еще не развита у ребенка 2-3 лет. Она появляется лишь к 3-4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
Нисходящие моторные системы
Высшие отделы головного мозга осуществляют свои влияния па деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути, которые группируют обычно в две основные нисходящие системы — пирамидную и экстрапирамидную.
В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения нервных окончаний в спинном мозге и функциональных различий, на следующие 2 системы: более молодую латеральную, волокна которой оканчиваются в боковых (латеральных) частях спинного мозга и связанную преимущественно с мускулатурой дистальных звеньев конечностей (сюда относят корково-спинномозговую и красноядерно-спинномозговую системы), и древнююмедиальную, волокна которой оканчиваются во внутренних (медиальных) частях белого вещества, связанную главным образом с мускулатурой туловища и проксимальных звеньев конечностей, состоящую из вестибуло-спинномозговой и ретикуло-спинномозговой систем.
Пирамидная системавыполняет 3 основные функции:
посылает мотонейропам спинного мозга импульсы — команды к движениям (пусковые влияния);
изменяет проведение нервных импульсов во вставочных спинальных нейронах, облегчая протекание нужных в данный момент спинномозговых рефлексов;
осуществляет контроль за потоками афферентных сигналов в нервные центры, выключая постороннюю информацию и обеспечивая обратные связи от работающих мышц.
Волокна пирамидной системы вызывают преимущественно возбуждение мотонейронов мышц-сгибателей, особенно влияя на отдельные мышцы и даже части мыши верхних конечностей, в частности на мышцы пальцев рук.
Экстрапирамидная системаоказывает обобщенные воздействия на позно-тоническиереакции организма от коры, мозжечка, промежуточного мозга и подкорковых ядер. Влияния этой системы передаются через корково-красноядерно-спинномозговой путь, составляющий функционально единое целое с «медленной» подсистемой пирамидного тракта, и через более древнюю медиальную систему (вестибуло-спинномозговую и ретикуло-спинномозговую системы).
Таким образом, среди нисходящих моторных систем, осуществляющих функцию контроля активности мотонейронов спинного мозга, можно выделить одну часть, которая обусловливает ф аз ну т двигательную деятельность — это «быстрая» подсистема пирамидного тракта, и другую часть — остальные нисходящие системы, которые обеспечивают регуляцию тонуса мышц и позных реакций организма. Из этих систем три системы обеспечивают повышение возбудимости мотонейронов мышц-сгибателей (корково-спинномозговая, корково-красноядерно-спинномозговая и коркоио-ретикуло-спинномозговая), а одна система (вестибуло-спинномозговая) — тормозит эти мотонейроны (рис. 1).
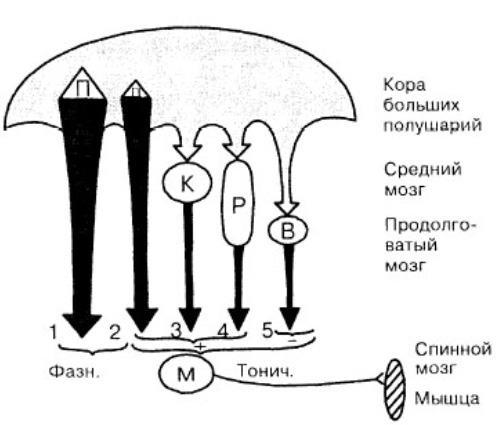
Рис. 1. Схема основных нисходящих путей регуляции двигательной деятельности
1 — быстрая подсистема и 2 — медленная подсистема корково-спиномозго-вого пути (пирамидного тракта); 3 — корково-красноядерно-спиномозговой путь. Латеральная система — I, 2, 3. Медиальная система — 4, 5. М — мотонейрон спинного мозга, получающий фазные (Фазн.) и тонические (Тонич.) возбуждающие (+) и тормозящие (-) влияния.